University of Southampton Falmouth Field Course 2013 26th June -
26th June 2013 -
Falmouth Tides (UTC): HW 07:20 5.1m
LW 13:50 0.3m
Cloud
cover: 100% -
Sea State: Flat -
Air Temperature: 14.9°C
Biological Discussion
Phytoplankton and Zooplankton
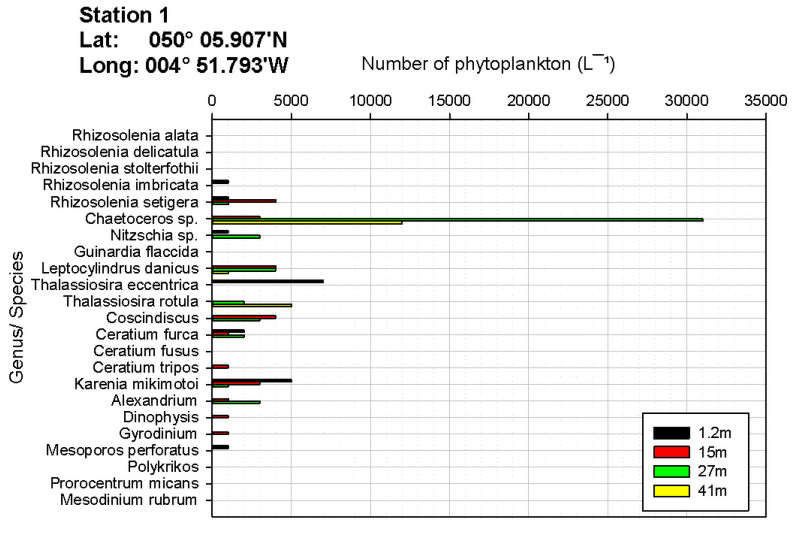
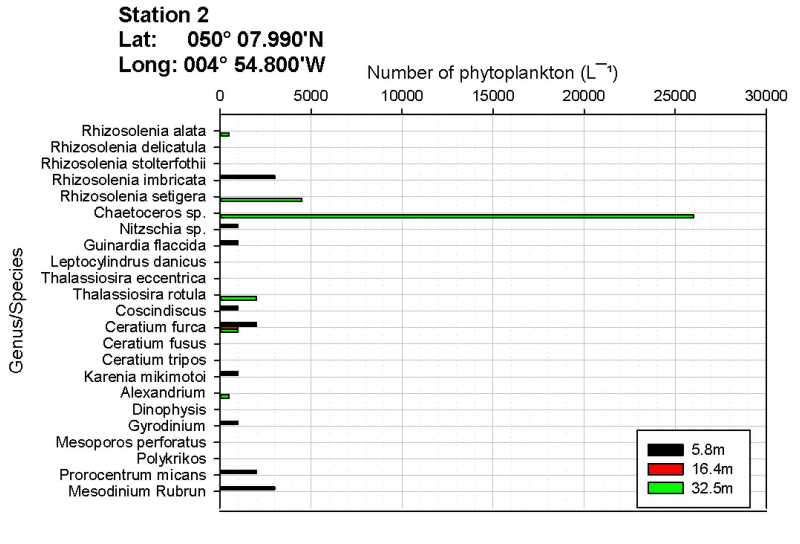
Edwards and John (1996) noted that planktonic abundance and distribution of species are influenced by different factors. Abundance is controlled by depth, tidal mixing and temperature stratification whereas the distribution of species is influenced by salinity, temperature, water inputs and the presence of local benthic and littoral communities. The number of phytoplankton in samples taken at varying depths alters as a result of mixing both vertically and horizontally. In most cases the chlorophyll maximum depth is the same as the maximum in phytoplankton biomass (Sharples, et al., 2001). This could be observed at station 1 where both the phytoplankton and chlorophyll maximum were observed at 27 metres. At station 2 the chlorophyll maximum has not been recorded due to a combination of a Niskin bottle failing to fire and a lost data point (chlorophyll).
The location of the maximum number of phytoplankton may change between the stations as a result of a change in the depth of the thermocline. This influences vertical variation in phytoplankton number in the water column. The location of the thermocline influences the mixing and movement of nutrients such as nitrate to shallower waters through upwelling; this in turn prompts phytoplankton growth at this depth (Keifer & Kremer, 1981). This can be observed at station 1 where the depth of the thermocline was 30 metres, which was approximately the same depth as the chlorophyll maxima (27 metres).
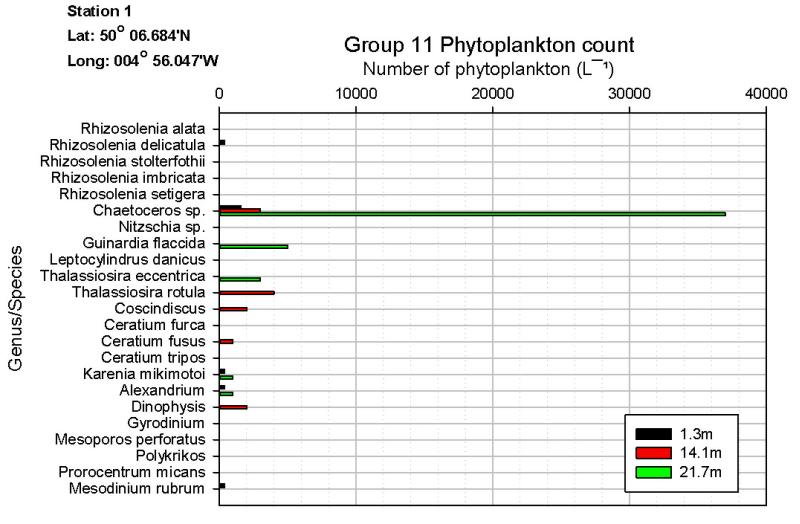
As shown on figures OB.4, OB.5, OB.6, the genus Chaetoceros is the most abundant phytoplankton at each station sampled (station 1 and 2 by Group 9, and station 1 by Group 11). Rees et al. (2009) measured a bloom of Chaetoceros debilis in 2007 which coincided with the UKs highest rainfall since records began. This is consistent with our data as the UK overall had 130% of the average rainfall in May this year (www.metoffice.gov.uk/climate). This leads to a prominent halocline, as seen in figure OP.5 on the physical results page, and may help the blooms of Chaetoceros spp. seen.
{kind=link}
{kind=link}
{kind=link}
Due to the lack of zooplankton nets deployed and therefore data collected over a
range of metres, it is difficult to compare the interactions between phytoplankton
and zooplankton. It can be suggested, however, that the relationship is not simply
predator-
Nitrate
Nitrate is an essential component for life in the ocean (Burford and Pearson, 1998), due to its use in proteins and nucleic acids. Smyth et al. (2010) noted that regions in the Western English Channel are often regulated by the availability of dissolved nitrogen. As seen in figure 10, station 1 shows an overall increase in nitrate however a decrease in phytoplankton. Officer and Ryther (1980) suggest this could be due to enrichment in nitrogen and phosphorus, which can be seen in figures OC.4 and OC.5 on the chemical results page, which shows nitrate to be a limiting factor. As previously mentioned, copepods control the abundance of ciliates (Turner, 2004), however it has also been shown that this indirectly regenerates ammonium by flagellates within the community, ultimately increasing the nitrate pool in the water column (Glibert et al. 1992).
Silicon
The concentration of silicon in the water column affects the number of diatoms, as
shown in figures OB.7, OB.8 and OB.9. Silicon is used by diatoms in the frustule
and is required for growth (Martin-
Figure OB.7 at station 1 shows a prolonged decrease in silicon concentration with increasing depth which is followed after a period of time by a decrease in the number of diatoms. Paasche (1973) states that the growth of phytoplankton in coastal waters will become seriosly limited by silicon only when most of the silicon originally present is removed due to a diatom bloom. This explains the time lag between a decrease in diatom numbers in the water column and decreased silicon concentrations.
An increase in diatom numbers with depth is also expected due to their high sinking
rates (Yool & Tyrrell, 2002). This is evident in station 2 (Group 9) and station
1 (Group 11) where the highest number of diatoms are found in depths below 20m (Station
1, Group 9 also increased diatom numbers between 0m-
References
Burford M. and Pearson, D. 1998, Effect of different nitrogen sources on phytoplankton
composition in aquaculture ponds, Aquatic Microbial Ecology, 15, 277-
Edwards, M. and John, A.W.G. 1996, ‘Chapter 4.3 Plankton’, Barne J.H., Robson, C.F., Kaznowska, S.S., Doody, J.P., Davidson, N.C. and Buck, A.L. (eds), Coasts and seas of the United Kingdom. Region 11 The Western Approaches: Falmouth Bay to Kenfig, Joint Nature Conservation Committee, Peterborough. (Coastal Directories Series).
Fenn, ME., Haeuber, R., Tonnesen, G.S., Baron, J.S., Grossman-
Glibert, P., Miller, C., Garside, C., Roman, M. and McManus, G. 1992, NH4+ regeneration
and grazing: interdependent processes in size-
Keifer, D. A., & Kremer, J. N. 1981, Origins of vertical patterns of phytoplankton
and nutrients in the temperate, open ocean: a strategic hypothesis. Deep-
Martin-
Met office official report, Climate summaries [online]. Available: http://www.metoffice.gov.uk/climate [accessed 2013, July 2nd].
Officer, C.B. and Ryther, J.H. 1980, The possible importance of silicon in marine
eutrophication, Marine Ecology Progress Series, 3, 83-
Paasche, E. (1973). Silicon and the ecology of marine plankton diatoms. I. Thalassiosira
pseudonana (Cyclotella nana) grown in a chemostat with silicate as limiting nutrient.
Marine Biology , 19 (2), 117-
Rees, A.P., Hope, S.B., Widdicombe, C.E., Dixon, J.L., Woodward, M.S. and Fitzsimons,
M.F. 2009, Alkaline phosphatase activity in the western English Channel: Elevations
induced by high summertime rainfall, Estuarine, Coastal and Shelf Science, 81, 4,
569-
Sharples, J., Moore, M. C., Rippeth, T. P., Holligan, P. M., Hydes, D. J., Fisher,
N. R. and Simpson, J. 2001, Phytoplankton distribution and survival in the thermocline.
The American Society of Lymnology and Oceanography , 46, 3, 486-
Smyth, T., Fishwick, J., Al-
Stoecker, D.K., Egloff, D.A. 1987. Predatation by Acartia tonsa Dana on planktonic
ciliates and rotifers. Journal of Experimental Marine Biology and Ecology, 110(1),
53-
Turner, J.T. 2004. The importance of small planktonic copepods and their roles in
pelagic marine food webs. Zoological Studies, 43(2), 255-
Yool, A., & Tyrrell, T. (2002). Role of diatoms in regulating the ocean’s silicon
cycle. Global Biogeochemical Cycles , 17 (4), 1-
Offshore

| Introduction |
| Methods |
| Results |
| Discussion |
| Physical |
| Chemical |
| Biological |
| Physical |
| Chemical |
| Biological |
| Introduction |
| Methods |
| Results |
| Discussion |
| Physical |
| Chemical |
| Biology |
| Physical |
| Chemical |
| Biology |