University of Southampton Falmouth Field Course 2013 26th June -
26th June 2013 -
Falmouth Tides (UTC): HW 07:20 5.1m
LW 13:50 0.3m
Cloud
cover: 100% -
Sea State: Flat -
Air Temperature: 14.9°C
Physical Discussion
Stratification Index (S)
A stratification index (S) equal to or higher than 3 represents highly stratified waters, an S value of –2 indicates highly turbulent waters and an S value equal to 1.5 represents a transition zone, thus providing optimum conditions for primary production (Simpson and Sharples, 2012). The S values for all stations (figure OP.1) are close to or higher than 3, which is indicative of stratified conditions within the vicinity of sampling stations. Overall, stratification index values (SI) at all stations show a well stratified water column within the sample area, where S values range between 3.923 (Station 1) and 2.757 (Station 2).
Temperature
The general trend for all stations is a decrease in temperature with depth (figure
OP.2). Mixing and the low temperature of the water column at Station 4 would be due
to the effect of surface wind mixing as well as tidal mixing, the influence of which
increases as the water depth decreases. The development of the observed seasonal
thermocline in the study area is typical of the coastal waters of south-
Salinity
Small range in salinity of 0.17 units between all four stations (figure OP.10) is to be exected, as station locations are offshore from Falmouth and thus the influnce of fresh water input from rivers discharging into Fal and Helford estuaries is not detected. The observed spike in the salinity profile at Station 2 at approximately 20 m depth (figure OP.8) is attributed to the mismaches in CTD sensors response time (mainly conductivity cell and temperature transmissometer). This corresponds to a spike at approximately 20 m seen in the density profile (figure OP.4), which is to be expected, as density is derived from salinity, temperature and pressure. The greater the salinity spike, the greater the lag in response time between the two sensors.
Figure OP.8: CTD derived vertical profiles of salinity at all stations showing salinity ‘spike’ at 20 m depth
Density
The depth of the pycnocline decreases from Station 1 to Station 2 and progressively disappears towards Station 3 and completely disappears at Station 4 (figure OP.4), where the effect of tidal and wind induced mixing increase and are more pronounced as the water depth decreases towards the mouth of the estuary. The developed pycnoclines at Stations 1 and 2 represent staticaly stable water columns, while waters at Stations 3 and 4 exibit turbulent character. This is in correspondence with the occurrence and change in flood and ebb tide flows (HW at 07:20 (UTC) and LW at 13:50 (UTC)).
Richardson Number (Ri)
Richardson number (Ri) is given by: Ri = N2/(du/dz)2, where N = Brunt-
For shallow estuarine and offshore environments the following simplifications could
be applied: pressure effect can be neglected, as well as g2/c2 term (g (m s-
Current velocity values for each station have been taken from the ADCP discharge summary table.
Figure OP.9: Vertical profiles of Richardson numbers at all stations
Overall, calculated Ri numbers at all stations (figure OP.9) fall below 0.25, indicating
that the flow at all four stations exhibits turbulent behaviour. It could be said
that that the Ri values in figure OP.9 do not correspond to the stratification index
values, which suggests that the water column between the stations is highly stratified.
However, flows are never truly laminar (Pickard and Emery, 1990), which can be seen
from the current velocity (m s-
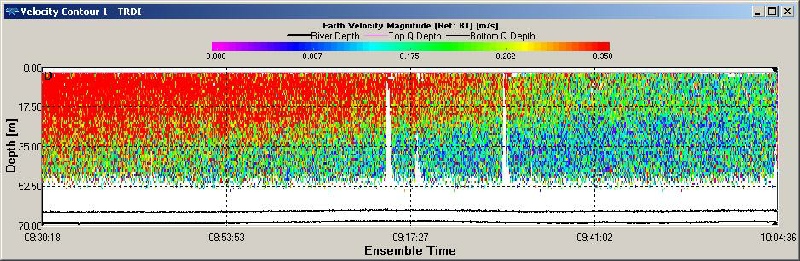
Figure OP.10 ADCP derived current velocity contour plot between Black Rock and Station 1
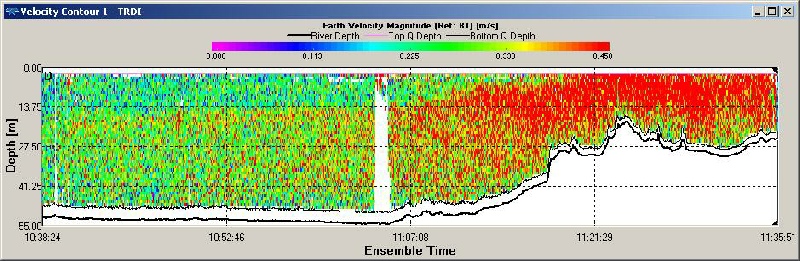
Figure OP.11 ADCP derived current velocity contour plot between Station 1 and Station 2
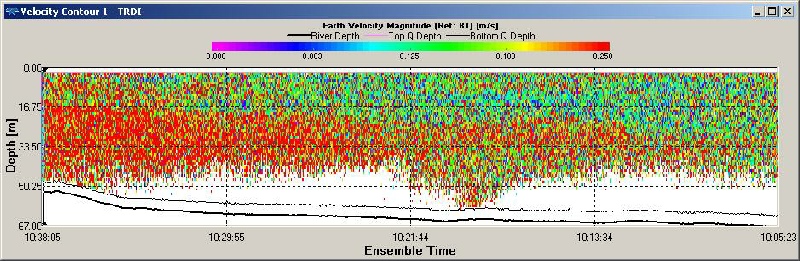
Figure OP.12 ADCP derived current velocity contour plot between Station 2 and Station 3
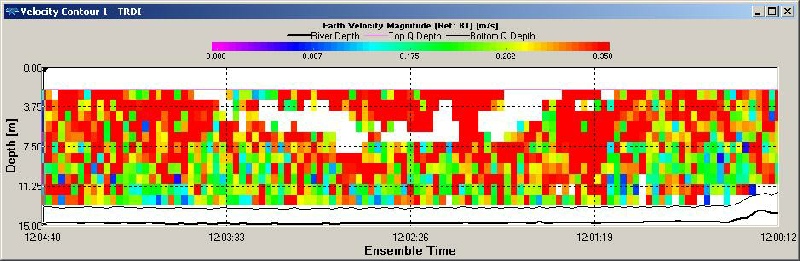
Figure OP.13 ADCP derived current velocity contour plot between Station 3 and Station 4
Fluorescence
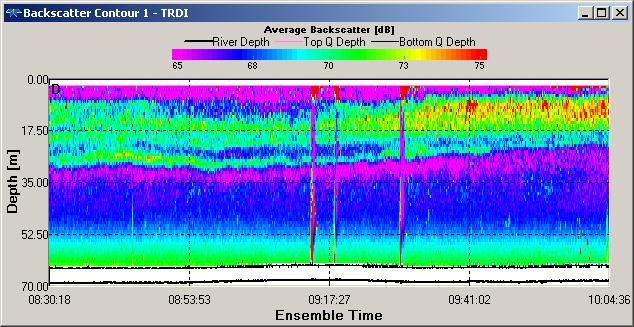
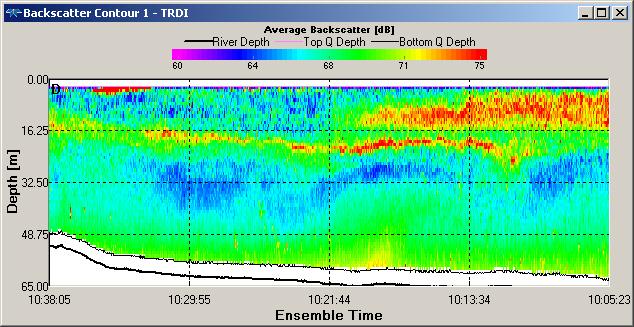
Backscatter contour plots for Station 1 and Station 2 (figure OP.14 & OP.15) show
the presence of zooplankton species between subsurface depths of 6m and 27.60m for
Station 1 and subsurface depths of 6m and 22.50m for Station 2, which indicates the
presence of zooankton. This corresponds to fluorescence peaks at ~ 30 m depth at
Station 1 (0.25 µg/ L) and at ~ 17 m at Station 2 (0.21 µg/ L), indicating that the
bottom of the euphotic zone (figure OP.6) provides optimum growth conditions for
zooplankton species at Station 1 and Station 2. Increased backscatter in both contour
plots (figure OP.14 & OP.15) is due to re-
Figure OP.14 ADCP derived backscatter plot between Black Rock and Station 1
Figure OP.15 ADCP derived backscatter plot between Station 1 and Station 2
Transmission
Transmission measures the percentage of light penetration through the water column
and is used as a proxy for the amount of suspended particulate matter (SPM) (Towns,
2010). At Stations 1 and 2 the profiles show highly variable transmission through
the water column (figure OP.7), due to stratified conditions (figure OP.2 & OP.4).
At Station 1 the sharp decrease in transmission at approximately 30m depth is most
likely due to a peak in phytoplankton numbers, which creates scattering within the
path-
Linking physical conditions with phytoplankton abundance and distribution within the water column
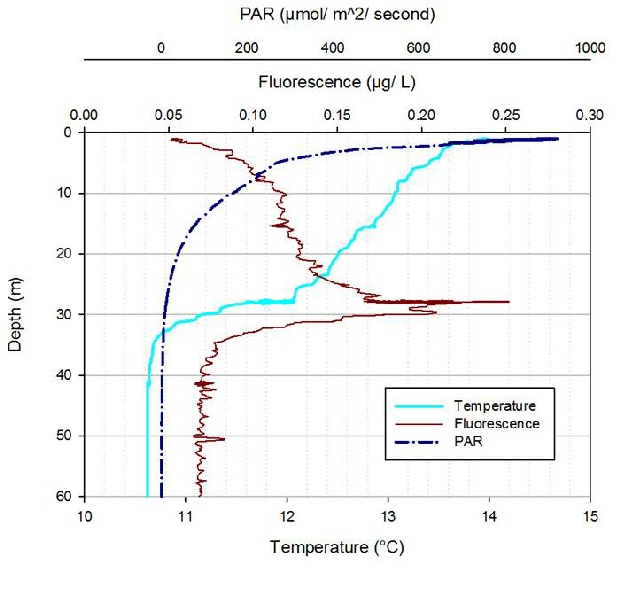
Figure OP.16: CTD derived vertical profiles of PAR, fluorescence and Temperature at Station 1
Combined vertical profiles of temperature, fluorescence and photosynthetic active
radiation (PAR) clearly show the effect of light availability and temperature on
phytoplankton species abundance (fluorescence vertical profile) and distribution
within the water column. Fluorescence is a measure of chlorophyll a (Chl a) concentrations
(µg/ L) and the observed peak at approximately 27.50m depth corresponds to the Chl
a maximum depth, which is supported by highest phytoplankton count of 3100 cellsL-
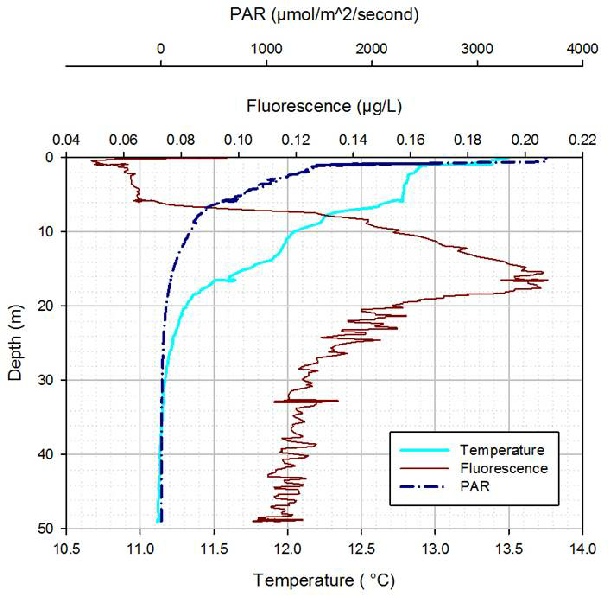
Figure OP.17: CTD derived vertical profiles of PAR, fluorescence and temperature at Station 2
Station 2 combined vertical profiles of temperature, fluorescence and PAR show the
same trend seen at Station 1 (figure OP.16), however, the thermocline is shallower
(between 15 and 20m). This results in the fluorescence peak occurring at this depth
range as well. However, the maximum count number of phytoplankton cells is 26000
cellsL-
References
Dyer, K., 1983, Mixing caused by lateral internal seiching within a partially mixed estuary, Estuarine and Coastal Shelf Science, Issue 26, pp. 51 – 66
Kirk, J., 2011, Light and Photosynthesis in Aquatic Ecosystems, Chapter 6.3: Downward
irradiance-
Pickard and Emery, 1990, Descriptive Physical Oceanography, Chippenham, Wiltshire, UK
Simpson, J. and Sharples, J. 2012, Introduction to the Physical and Biological Oceanography of Shelf Seas, , Cambridge University Press, Cambridge, UK
Smyth, T., Fishwick, J., Al-
Towns, J. 2010, Relationship of light transmission, stratification and fluorescence
in the hypoxic region of Texas-
Viljanen, M. Holopainen, A-
Offshore
-1.JPG)
.JPG)

| Introduction |
| Methods |
| Results |
| Discussion |
| Physical |
| Chemical |
| Biological |
| Physical |
| Chemical |
| Biological |
| Introduction |
| Methods |
| Results |
| Discussion |
| Physical |
| Chemical |
| Biology |
| Physical |
| Chemical |
| Biology |