Biogeochemical Impacts
Emiliania huxleyi blooms may be very important in terms of global
planetary temperature. When these blooms appear over large expanses of the
ocean they have myriad effects on the water and on the atmosphere above.
Although each cell is invisibly small, there can be as many as a thousand
billion billion (10^21) of them in a large bloom, and the population as a
whole has an enormous impact. Ongoing work is trying first of all to
estimate whether the net effect of the blooms is to exacerbate or to
ameliorate global warming; subsequently the magnitude of the combined
forcing on global temperature will be estimated.
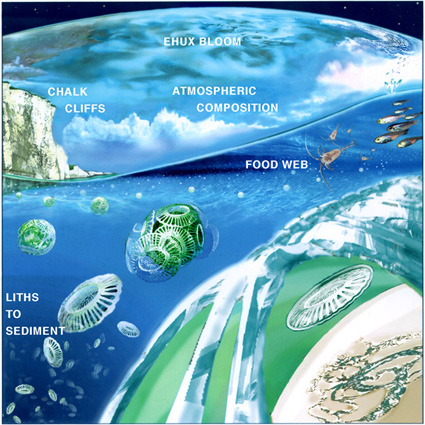
Ehux blooms are processed through the food web, with viruses, bacteria
and zooplankton all contributing to the demise and decomposition of blooms.
Some debris from the bloom survives to sink to the ocean floor, taking
chemicals out of the water column. While they live and when they die, the
phytoplankton cells leak chemicals into the water. A bloom can be thought
of as a massive chemical factory, extracting dissolved carbon dioxide,
nitrate, phosphate, etc from the water, and at the same time injecting other
chemicals such as oxygen, ammonia, DMS and other dissolved organic compounds
into the water. At the same time, the chemical factory pumps large volumes
of organic matter and calcium carbonate into the deep ocean and to the ocean
floor. Some of this calcium carbonate eventually ends up as chalk or
limestone marine sedimentary rocks, perhaps to cycle through the Earth's
crust and to reappear millions of years later as mountains, hills and
cliffs. Picture courtesy of
Glynn Gorick
(copyright held).
Global abundance: a
global satellite study
by Brown & Yoder detected an annual area of blooms of 1.4 million km2. This
is the total area each year that was classified as coccolithophore blooms,
i.e. that was unobscured by clouds and that satisfied the imposed thresholds
in terms of brightness, size of bloom area, etc. We know that there is also
a lot of coccolithophore productivity in non-bloom areas, for instance in
oligotrophic tropical oceans (Balch & Kirkpatrick, 1996), where there is a
continual background turnover of coccolithophores even though their
abundance never reaches bloom proportions.
Total calcite production: because calcite fluxes are not usually
determined just for Ehux calcite, we will consider the biogeochemical
impacts of global ocean calcite productivity, i.e. productivity of
Ehux and other coccolithophores, and also other organisms. Although
Ehux is frequently the most abundant coccolithophore (by number) in
seawater samples, because of its smaller size it doesn't always dominate the
calcite flux (by weight). Other coccolithophores with larger coccoliths,
such as Calcidiscus leptoporus will usually be more important to the
calcite export flux. From productivity estimates, sediment trap
measurements, and from geochemical models, we can estimate how much calcium
carbonate (calcite) is produced annually by coccolithophores and
foraminifera and coral reefs (the main producers of calcite in the oceans).
Several authors (Morse & Mackenzie, 1990; Wollast, 1994; Milliman, 1993;
Shaffer, 1993; Archer & Maier-Reimer, 1994) have estimated total global
calcite productivity as 0.63, 1.13, 0.64, 1-2 and 1.2 (average ~1.0) Gt
calcite-C year-1. The relative contributions of coccolithophores,
foraminifera and other organisms to the total remain relatively poorly
known.
Total calcite burial: a significant percentage of the produced
calcite (somewhere between 0 and 80%) probably gets dissolved before
incorporation into sediments, which raises the question: how much calcite
does get buried? This figure is more easily obtained, from averaging the
amount of calcite in different marine sediment cores and dividing by the
rate of accumulation of the material in the cores. Milliman (1993)
estimates a calcium carbonate sedimentary accumulation rate of 0.46 Gt C
year-1.
A lot of the dissolution of calcite (the difference between production and
burial) probably occurs in surface waters, but also a lot occurs at very
great depths. From comparison of the amounts of calcite in different marine
sediment cores, it is apparent that no calcite at all reaches some parts of
the sea bed, especially the sea bed underneath the very deepest waters.
This because of what is known as the calcite lysocline (Broecker &
Peng, 1982). Below a certain depth (4-5km or so) the great pressures cause a
change in the seawater chemistry such that calcite is forced into solution.
The water suddenly becomes much more corrosive to calcite, and all of the
calcite fragments raining towards the sea floor suddenly get rapidly
dissolved below the critical lysocline depth.
Types of climate forcing: there are several ways in which
coccolithophores influence regional and global temperature:
1. Ocean Albedo: as described on the
optical impacts page,
coccoliths act somewhat like little mirrors, because of the special optical
properties of calcite. This causes a typical bloom (containing 100 mg m-3
of calcite carbon) to increase the ocean albedo from ~7.5% to ~9.7%, as
shown in the model-derived diagrams below. Another type of optical model
(more accurate for this purpose) calculates an increase from ~6.2% to ~9.7%.
If each bloom is assumed to persist for about a month, then an annual
coverage of 1.4 x 10^6 km squared (see above) will increase the global
annual average planetary albedo by
(9.7-6.2) x (1/12) x (1.4/510) = 0.001%
where 510 is the surface area of the Earth in 10^6 km squared. This is a
lower bound on the total impact, because sub-bloom concentration coccolith
light scattering will have an impact, over much larger areas (estimated
maximum albedo impact = 0.21%), as will cloud-obscured blooms, which were
not included in the global total of Brown & Yoder because they could not be
seen by the satellite.
A 0.001% albedo change corresponds to a 0.002 W m-2 reduction in incoming
solar energy, whereas an albedo change of 0.21% causes a reduction of 0.35 W
m-2. These two numbers can be compared to the forcing due to anthropogenic
addition of CO2 since the 1700's, estimated to be about 2.5 W m-2.
Coccolith light scattering is therefore a factor of only secondary
importance in the radiative budget of the Earth.
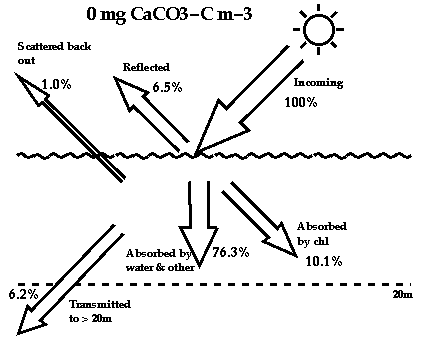
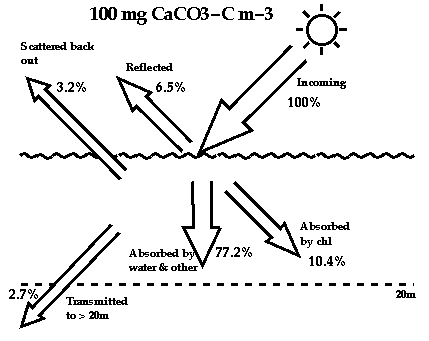
Photon budgets for water without any coccoliths (top diagram), and for
water with a concentration of 100 mg m-3 of calcite carbon in the form of
coccoliths.
2. Ocean Heat Retention: the scattering caused by coccoliths causes
more heat and light than usual to be pushed back into the atmosphere. It
also causes more of the remaining heat to be trapped near to the ocean
surface, and only allows a much smaller fraction of the total heat to
penetrate to deeper in the water (see photon budgets above). Because it is
the near-surface water which exchanges heat with the atmosphere, all three
of the effects just described conspire to mean that coccolithophore blooms
may tend to make the overall water column dramatically cooler over an
extended period, even though this may initially be masked by a warming of
the surface skin of the ocean (the top few metres). The importance of this
effect, both regionally and globally, is currently being worked upon but has
not yet been established.
3. Cloud Albedo: phytoplankton produce a sulphur compound called
dimethyl sulphide (DMS), with coccolithophores and other species producing
100 times more of it than diatoms (Keller, 1989). After the cells die, and
after several subsequent biological and chemical transformations in the
ocean and in the atmosphere, some of this DMS eventually ends up as cloud
condensation nuclei (CCN) in the atmosphere. These CCN help to stimulate
cloud formation, especially in areas where other sources of CCN are in short
supply, and the amount of CCN is limiting possible cloud growth. In this
way, coccolithophore productivity helps to increase the reflectiveness
(albedo) of the planet by allowing more clouds to form. Due to the
complexities of the processes involved in the transformation from
phytoplankton DMS to CCN, we do not yet have a good estimate of how
important this forcing is.
4. CO2 Greenhouse: all phytoplankton growth removes carbon dioxide
(CO2) into organic matter and reduces atm. CO2 (click
here
for explanation). However, coccolithophores are unique in that they also
take up bicarbonate (HCO3), with which to form the calcium carbonate of
their coccoliths. The chemical reaction for coccolith formation is:
Ca + 2HCO3 ---> CaCO3 + H2O + CO2
There are three forms of dissolved carbon in seawater: CO2, HCO3 and CO3;
and carbon can shift very easily from being in one of these dissolved forms
to being in another. How much of the total carbon is in each form is
determined mainly by the alkalinity and by the water temperature. When the
seawater carbon system is perturbed by coccolithophore cells removing HCO3
to form coccoliths, this causes a re-arrangement of how much carbon is in
each dissolved form, and this rearrangement takes place more or less
instantaneously. The removal of 2 HCO3 molecules and the addition of one
CO2 molecule changes the alkalinity and this indirectly causes more of the
dissolved carbon to be pushed into the CO2 form. Although the total
dissolved carbon is obviously reduced by removal of dissolved carbon
(bicarbonate ions) into solid calcium carbonate, yet the total effect,
paradoxically, is to produce more dissolved CO2 in the water. In this way,
coccolithophore blooms tend to exacerbate global warming (by causing
increased atmospheric CO2), rather than to ameliorate it, as is the case
when dissolved CO2 goes into new organic biomass.
However, recent work is showing that additional properties of coccoliths may
make the situation yet more complicated. Coccolith calcite is rather dense
(2.7 kg per litre compared to seawater density of 1.024 kg per litre), and the
presence of coccoliths in zooplankton faecal pellets and `marine snow' (the
two main forms in which biogenic matter sinks to the deep ocean) causes them
to sink more rapidly. Slow-sinking organic matter may also adhere to the
surfaces of coccoliths, hitching a fast ride out of the surface waters.
If organic matter sinks faster then there is less time for it to be attacked
by bacteria and so more of the locked-in carbon will be able to escape from
the surface waters, depleting the surface CO2.
A recent paper (Buitenhuis et al, 1996) argues that this co-transport
of organic matter with coccoliths offsets the atmospheric CO2 increase that
would otherwise be caused, and makes coccolithophore blooms act to oppose
global warming, rather than to intensify it.
References
Ehux
home page
Toby Tyrrell : T.Tyrrell@noc.soton.ac.uk