![]()
Eleanor Colling, Debbie Hembury, Jeremy Lynas, Charlotte Main, Dawn Morgan, Luke O'Mara, Lewis Rowntree, Edward Smith, Edmund Willy.
Contents

Figure 1 Members of group 11 aboard the Terschelling
Plymouth Sound, England, is located south of the confluences of the rivers Tamar and Lyhner and of the Tamar and Tavy, Devon. This web page summarises the preliminary findings from a group (fig. 1) of nine people, working in conjunction with eleven other groups, during an extended sampling effort carried out in Plymouth Sound and associated estuaries during June and July 2004. The aims of the investigation were to study the physical, chemical and biological characteristics of the estuary, and to draw appropriate conclusions about the processes governing these characteristics. Investigation by the groups as a whole extended from riverine end members near Calstock on the River Tamar out as far as E1, beyond Eddystone Rocks, offshore.
The estuarine area investigated has a high diversity of habitats with a wide variety of flora and fauna. The northern shore of Plymouth Sound features a reef of submerged Devonian limestone. The Tamar Estuary is partially mixed and mesotidal, with a tidal range of 2.2-4.4m on neaps and 0.8-5.5m on springs relative to Chart Datum. There are extensive tidal mudflats and salt marshes. The mudflats are some of the largest in the South West, and provide habitats for an abundance of invertebrate species.

A brief study of the geological composition and fold geometry of an area of Heybrook Bay was undertaken on 24/06/2004. Strata layers that have been compressed and consequently folded are evident throughout Renney Rocks, which are located at the southern end of Heybrook Bay. Ancient plate movements and continental drift created sufficient stresses and pressures to create a series of fractures that radiate around
a fault line. The compressive stress during formation of this geological feature resulted in a right lateral dextral fault with major displacement visible along the fault line. A conjugate set of fractures can be seen orientated at between 30º - 45º to the fault. Diagrams supporting this study Fig 2. View of
are in the group log book. Renney Rocks
A section of cliff face that runs parallel to the Renney Rocks area gives an indication of ancient deposition events, including periods of higher energy deposition. The cliff face has a reddish colour, indicating the presence of oxidised iron. This is characteristic of warm, humid climate periods at the time of deposition. The darker red band that is dominant throughout the cliff section demonstrates a period of highest incident radiation, evidently at the most recent climatic optimum (the Holocene) around 8000 years ago. This band consists of unsorted clasts imbedded within a coarse matrix and is representative of a "pulse" of energy, such as a landslide or mudslide during deposition. Throughout the strata within the cliff section the presence of small high-energy deposit layers indicate further, sudden "pulse" events, adequate to form stratified sub layers. The angularity of the clasts that are visible along the cliff section indicate that weathering processes dominated by frost shattering were probably evident at that depositional period. Between 10000-18000 years ago the study area was predominantly periglacial, which accounts for the occurrence of weathering processes such as freeze-thaw.
The aim of this
fieldwork was to investigate the chemical and biological characteristics of the
River Tamar from the riverine end member (0 salinity) found near Calstock (site
0: 50°29.616'N 004°11.684'W), to
higher salinities near to the Tamar Bridge (site 30: 50°24.555'N 004°12.207'W)
by using two small boats. Data
collected on the day (02/07/04) were
shared with group 10, who were working on the Bill Conway further down the
estuary. An overlap in salinities sampled by the two groups was aimed for, in
order to provide a continuum of measurements from the riverine end member to the
saline end member. Surface water samples were analysed for concentration of nitrates,
phosphates, silicate and chlorophyll. One further surface water sample per boat was taken using a Niskin bottle. This
sample was analysed for concentration of dissolved oxygen analysis. At each sampling
station, multi-parameter probes were used to measure salinity, temperature,
dissolved 02, pH and depth in a vertical profile.
The small boats (fig. 3) started up-river of Calstock at close to zero salinity. Samples were taken moving downriver at salinity intervals of 2, staggered between the two boats - each boat stopped at a station with every rise in salinity by 4. It was necessary to work in a direction moving downriver because of the 11:26 GMT high water at Calstock. Shallow depths at low water (0m above Chart Datum at low water) meant that
starting at Calstock in this way allowed a greater salinity range to be sampled in the river.
Three zooplankton tows at three separate salinity stations were carried out in order to measure abundance of zooplankton. The zooplankton nets were deployed by mooring them to a buoy at the chosen sites so that the river current was able to flow into the net itself. Eulerian style sampling was carried out, by reading a flowmeter attached to the zooplankton nets.
Figure 3 . Action on the boat
Phytoplankton samples were obtained at the same sites as the zooplankton tows. Phytoplankton samples were preserved with iodide so that counts could later be performed in the laboratory. Both zooplankton and phytoplankton samples were used to give an indication of the relative abundance of these groups at different salinities in the river, although the small (3) number of samples taken precludes many specific conclusions from being made about biological processes.
Water densities were calculated from temperature and salinity data from the multi-parameter probes used at on the small boats. This was done for every station sampled during the ribs boat work, between station 0 (GPS 50°29.616N, 004°11.680W, time 11.25GMT) and station 32 (GPS 50°23.978, 004°12.540, 14.57GMT). These densities are displayed on the contour maps (figures 4 and 5). Fig 4 shows density change with depth and salinity which, as would be expected from the equation of state, increases with increasing salinity. There is little change in density with depth at constant salinity, demonstrating that at these shallow depths pressure is insufficient to have any significant effect on density.
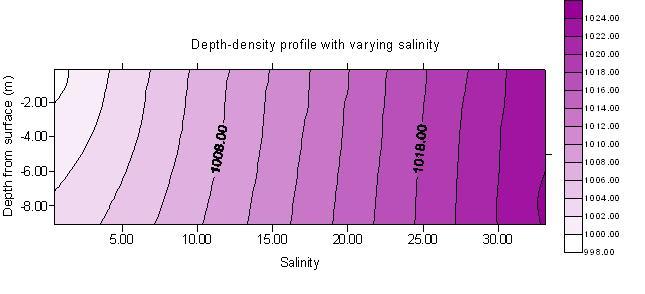
Figure 4. Density depth profile with varying salinity
Fig 5 shows the change in density with depth and station number. This provides a plot of the density stratification progressing downstream – a longitudinal cross section of the water body - showing that density increases with depth and site number (and therefore distance downstream). This demonstrates the gradient of mixing of less dense fresh water from the head of the estuary – which lies closest to the surface - with more saline, denser water at the marine end of the estuary. The saline water mixes with the overlying less dense fresh water, as can be seen in the density-depth-site number plot (fig 5).
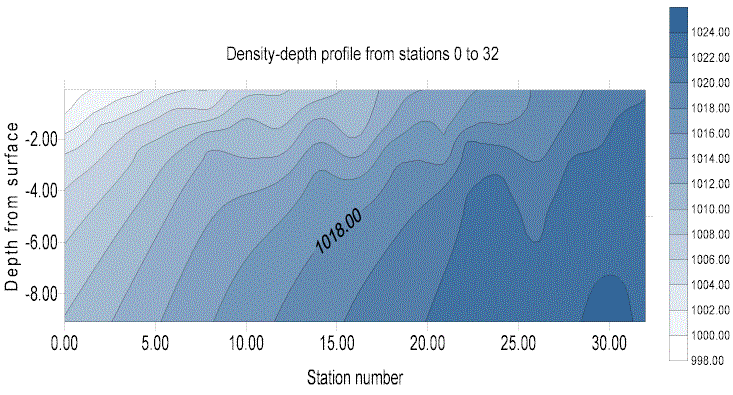
Figure 5. density depth profile from sites 0-32.
These data are limited by the fact that the data were taken at varying tidal states. The data would need to be taken simultaneously to produce an absolutely accurate plot of the density stratification of the water column. These data do, however, show the trends that would be expected from the mixing mechanisms of an estuary.
Analysis of temperature vs. salinity depth profiles for sites 0-32.
Data from group 11 estuarine sampling effort using small boats can be found in the following directory:
group11\all_work\25-06rib\rib-data
For the ribs work, most site numbers correlate approximately to the surface salinity measured at that sampling station.
At Calstock (at 0 surface salinity) the T/S profiles show fresh river water overlying colder saline water. There are a shallow thermocline and halocline at approx. 1m depth, and a temperature minimum of 15.6°C (at the bottom).
At sites 4, 6 & 8 there is a general increase in salinity with depth and a decrease in temperature with depth (as seen in all T/S profiles collected). The thermocline is still shallow at 1m depth.
At sites 10, 12 & 14 the thermocline is still at 1m depth, the salinity still increases with depth and there is a steady decrease in temperature deeper than the 1m thermocline.
At sites 16 and 18 the thermocline is deeper (2m depth). The structure of the temperature remains roughly the same with approximately the same decrease in temperature with depth.
At sites 22, 24 and 26 the water column is deeper and there is a further decrease in temperature with depth. The surface salinity increases from 20 to 23.
At sites 30 & 32 (by the Tamar Bridge) the surface salinity reaches 30 and increases still with depth to 32.7. The structure of the temperature profile remains approximately the same.
Analysis of salinity vs. euphotic zone depth
Euphotic zone depth was calculated as Secchi disk depth multiplied by 3. With increasing salinity, the euphotic zone depth was seen to increase. The magnitude of increase in euphotic zone depth between salinity intervals increased with proximity to the saline source.
As salinity increases, so too does the concentration of Na+ ions. These ions encourage suspended particulate matter (SPM) in the water column to coalesce, leading to the formation of larger colloidal particles. These particles have a greater tendency to fall out of suspension to the seabed, therefore increasing light penetration and deepening the euphotic zone.
Chemical Analysis.
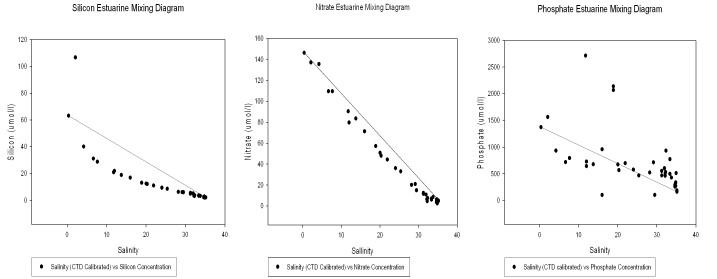
Fig 6: A (left), B (middle), C (right). Estuarine mixing diagrams for Silicate (A), Nitrate (B) and Phosphate (C) in the Tamar estuary.
Silicon
Since all measured silicate concentrations (except one) fall below the theoretical dilution line (TDL), non-conservative removal of silicon was observed (fig 6 A). One anomalous result was recorded at site 2 (at Calstock). This value was unusually high possibly due to the influence of a small tributary in close proximity to the site and up river.
Nitrate
Most data points occur in a moderately straight line slightly below the TDL (fig 6 B). This supports findings by Morris et al., (1981) who found an essentially conservative relationship between nitrate and salinity in the upper reaches of the Tamar Estuary. There appear to be no anomalous recordings for nitrate.
Phosphate (Data from Ribs only)
Phosphate shows a non-conservative relationship in the estuary, with data points occurring both below the TDL (fig 6 C), indicating removal (lower salinities) and also above the TDL (higher salinities). At salinities above 25 there is an influx of phosphate into the estuary, shown by points above the TDL. This may be due to geographical features close to these sites. It is possible that the steep sides of the surrounding banks, large areas of agricultural land and increased surface run-off due to recent increased precipitation events, have elevated the phosphate concentration at these sites. Non-conservative and non-biological removal of phosphate could also be as a result of chemical precipitation of dissolved phosphate at lower salinities, as found in the Tamar Estuary by Morris et al., (1981).
Chlorophyll 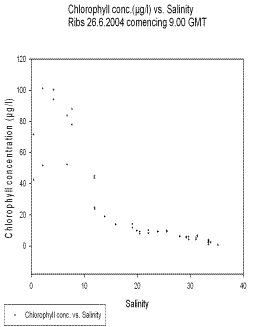
The concentration of chlorophyll in the estuary is higher at low salinities (see fig 7), indicating that there is a decreasing abundance of phytoplankton heading seawards. This can be explained by increased grazing pressure by zooplankton further to seaward. Though species specific, zooplankton tolerance of low salinities may be less than that of phytoplankton in many cases. This may allow phytoplankton to increase in abundance in the nutrient-rich waters further up the estuary in the absence of heavy grazing pressure by zooplankton. A Secchi depth of 19cm at the lowest salinity sampled (site 0) would allow enough light in the top 50cm (approx.) of the water column, to support primary production. Two zooplankton counts carried out in the estuary indicate that the abundance of zooplankton increases heading seawards, though more comprehensive studies are required to form definite conclusions. Higher primary production in the water column would be expected towards the seaward end of the estuary, because of increased euphotic depth. This would support a larger population of zooplankton at higher salinities. These zooplankton would be able to exploit a food source of phytoplankton to a limited extent up the estuary.
The phytoplankton population seen at the marine end of the estuary exhibit features of the end of a spring bloom, being dominated by diatoms. As silicate is undergoing removal at the marine end of the estuary, diatoms
may be becoming limited by silicate concentration. The Tamar Estuary has been shown to exhibit strong turbidity maximum in its low-salinity reaches (Grabemann et al., 1997). This results in a shallow euphotic depth at low salinities. Despite this, phytoplankton are able to exploit the nutrient-rich (figs 6 A, B, C) waters found at low salinities.
Fig 7. chlorophyll concentration vs salinity
Dissolved Oxygen
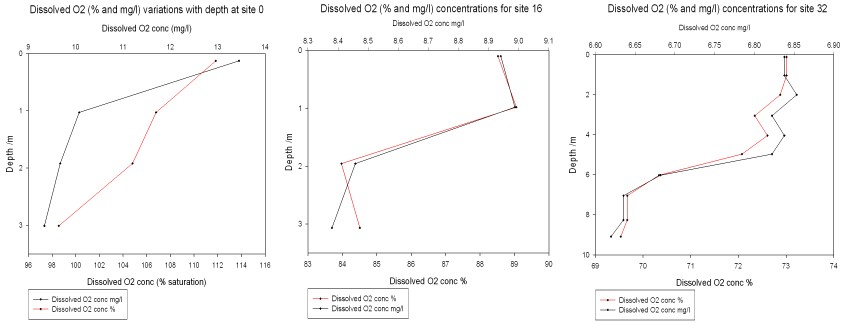
The
dissolved oxygen depth profiles show in general a decrease in concentration with
depth. The graphs above show dissolved oxygen depth profiles for close to zero
salinity, mid-range salinity and high salinity (~32) values. Oxygen
concentration in both % saturation and mg/l is observed as decreasing with
increasing salinity. The
chemical reactions of photosynthesis evolve oxygen. Therefore higher
concentrations of dissolved oxygen are to be expected in the surface waters
where phytoplankton are actively photosynthesising.
The approximate euphotic depths calculated from the Secchi disk
measurements at each station are 0.57m for station 0, 1.92m for station 16 and
4.8m for station 32. At each station,
dissolved oxygen concentrations decrease rapidly around and below the
corresponding euphotic zone depth, showing that some of the oxygen present in
the surface waters is likely to have been evolved by phytoplankton.
It should be noted that oxygen levels in surface waters will also be
elevated by physical processes, e.g. diffusion and wave action, and the presence
of dissolved oxygen is not entirely attributable to biological activity.
During the summer months the waters offshore
from
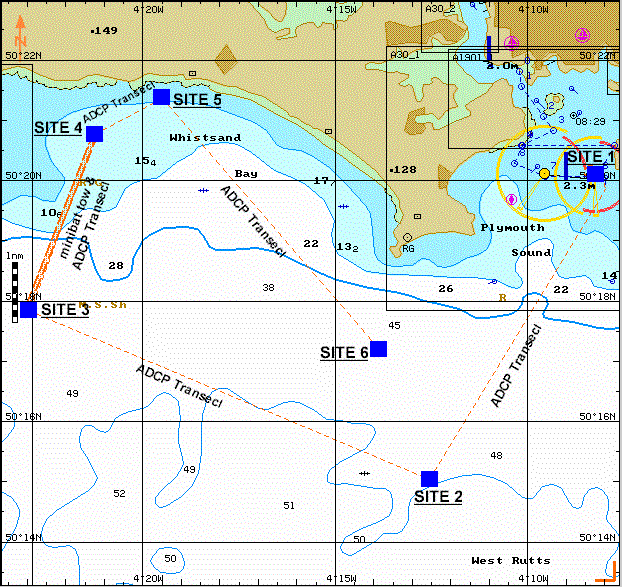
Figure 9. Map showing sites sampled (water samples and CTD), ADCP transects and miniBAT transect.
Six stations were sampled in the water off the
coast of
| ADCP transect time (GMT) | Start position | End position | Location |
| 08:30 - 08:38 | 50º 21.900'N 004º 07.900'W | 50º 20.152'N 004º 08.375'W | dock to site 1 |
| 09:10 - 09:55 | 50º 20.152'N 004º 08.375'W | 50º 15.062'N 004º 12.571'W | site 1 to site 2 |
| 09:55 - 10:20 | 50º 15.062'N 004º 12.571'W | 50º 15.062'N 004º 12.571'W | stationary at site 2 |
| 10:49 - 12:02 | 50º 15.062'N 004º 12.571'W | 50º 17.823'N 004º 22.950'W | site 2 to site 3 |
| 12:02 - 12:40 | 50º 17.823'N 004º 22.950'W | 50º 17.823'N 004º 22.950'W | stationary at site 3 |
| 12:40 - 13:33 | 50º 17.823'N 004º 22.950'W | 50º 20.778'N 004º 21.186'W | site 3 to site 4 (onshore transect) |
| 13:55 - 14:46 | 50º 21.395'N 004º 19.490'W | 50º 17.216'N 004º 13.876'W | site 5 to site 6 |
| 14:46 - 15:36 | 50º 17.216'N 004º 13.876'W | 50º 17.216'N 004º 13.876'W | stationary at site 6 |
Table 1. Positions and transect times for offshore ADCP.
Data from ADCP transects can be linked with
those from the CTD dips (at the start and end of transects) and also zooplankton hauls, in order to provide a
holistic interpretation. (Note: printouts for all CTD dips and ADCP transects
for the offshore boat work are in the group logbook).
The CTD profile at site
1 (behind the breakwater - see table 2 for positions of all CTD sites) shows
a thermocline and a halocline that are both steep and shallow (2-3m). There is a
0.5°C change in temperature across the thermocline and a change in salinity of
approx. 1 across the halocline. Fluorescence readings are low at the surface,
and higher beneath the thermocline. This could represent phytoplankton becoming
trapped beneath the thermocline during a period of stormy weather (gales and
heavy rain) five days previous to sampling, followed by some calmer weather
during the day before. Strong winds would have mixed the ~12m deep water column,
redistributing phytoplankton down throughout. During the ensuing calm weather, a
large input of fresh river water could have contributed to the re-stratification
of the water column, with the result that many phytoplankton could have remained
trapped at depth.
| Site number | Position | Time (GMT) |
| 1 | 50º 20.152'N 004º 08.375'W | 08:38 |
| 2 | 50º 20.062'N 004º 12.571'W | 09:55 |
| 3 | 50º 17.823'N 004º 22.950'W | 12:02 |
| 4 | 50º 20.778'N 004º 21.186'W | 13:33 |
| 5 | 50º 21.395'N 004º 19.490'W | 13:55 |
Table 2. Positions of CTD dips
Moving offshore from site 1, the ADCP
transect (fig. 10 data file in directory X:\group11\all_work\28_06off\off_data\off_raw\adcp)
shows the inshore turbid waters as uniformly red on the left side of the image.
As the transect moves offshore (looking from left to right) there is a layer of
backscatter that is shallow (just below the surface) to start with; then
deepening to approx. 8m. This scattering layer is probably the chlorophyll
maximum. The chlorophyll maximum grows deeper moving offshore because the water
column is stabilised by solar heating during the summer, producing a deeper
thermocline. Phytoplankton generally grow optimally at the depth of the
thermocline because this is usually the depth where there is optimum
availability of both light and nutrients. Offshore waters off
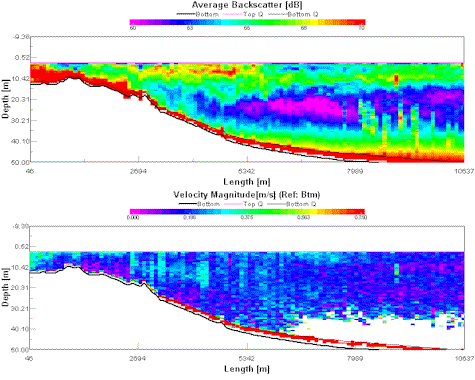
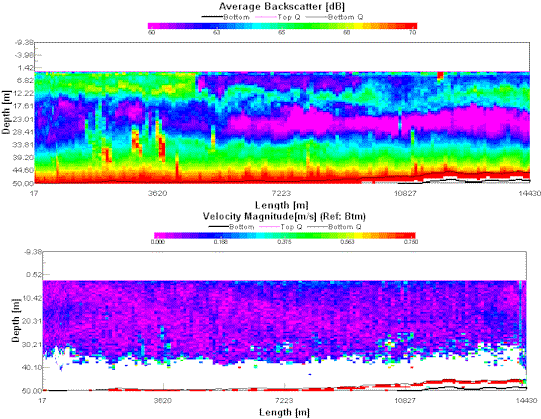
Figure 10 (left) ADCP transect between sites 1 and 2 showing the deepening scattered layer . Figure 11 (right) ADCP Transect between sites 2 and 3 showing the observed front
The CTD profile at site 2 shows a
thermocline that is less steep, extending over a greater depth. Note: the
thermocline at this site (site 2) represents a change in temperature of approx.
2 °C, from 14 °C down to 12 °C. At site 1 the change in temperature across
the thermocline was only approx 0.5 °C. Since site 1 is further inshore and
behind the breakwater, the warmer river water would have a greater influence
here. Offshore, at site 2, there is colder (12 °C) water at depth. The river
water from the estuary does not have a direct effect. Surface water is warmed by
solar heating and the water column hence becomes stratified. The effect is seen
over a greater distance vertically: the thermocline extends from near the
surface to approx 40m depth, and is gradual.
The ADCP data from the transect between sites 2 and 3 show a frontal system (fig. 10). This can be seen in the middle of the image as a thick scattered layer from ~14m depth extending up to near the surface (green) meeting a layer with low scattering (blue) this second layer overlies the scattered layer of the first water mass. The velocity magnitude plot does not show any specific anomalies, and it is likely that the front is being passively moved by currents in the water.
The miniBAT was towed from just after the start of the transect from site 3 (start GPS 50°18.098N 004°21.186W, time 12.55GMT) for part of the transect distance to site 4, stopping at 50°20.778N 004°21.186W, 13.33GMT. The depth-time profiles for several parameters are displayed below, dark green representing the higher values and blue the lower values in each case. The path of the miniBAT through the water is also marked on each plot with a light green track plot. The path is spread well throughout the body of water sampled, so the data can be expected to be accurate, perhaps with the exception of the data below 15m depth for time 0.00-20.00 (arbitrary time units) for which there are no real data.
Soon after beginning of the tow an there is an area of increased fluorescence, indicating presence of chlorophyll α, at a depth of approximately 15m. This correlates with an observed decrease in transmittance in the same area, suggesting presence of phytoplankton biological activity. The euphotic depth calculated from the Secchi disk measurements at site 3 was 25.5m, confirming that light-dependent biological activity could be present at 15m depth.The transmittance plot shows decreased transmittance towards the end of the tow, between the surface and 8m depth. This is not coupled with an increase in fluorescence, indicating that the increased transmittance is due to the presence of suspended particulate matter rather than biological activity. Being closer to the shoreline, it is possible to suggest that this is caused in turn by an increase in wave action and mixing processes in shallower water due to shear stress with the sea bed. Wave action would also increase the amount of oxygen in an area; this is supported in the dissolved oxygen data, with an increase in the amount of oxygen in the same area. However, it should be noted that the column of very low values just before this section is present throughout the water column and is likely to represent a failure in the instrumentation resulting in erroneous data.
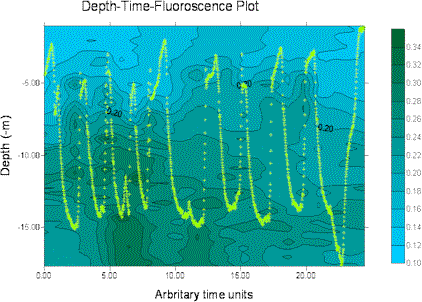
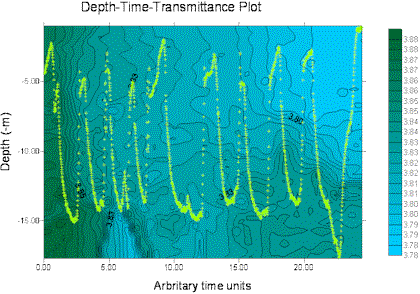
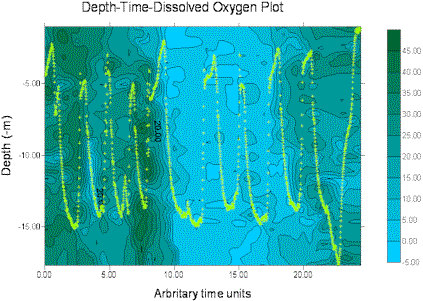
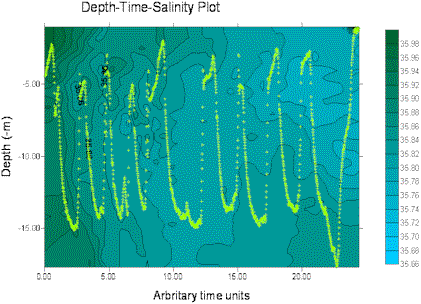
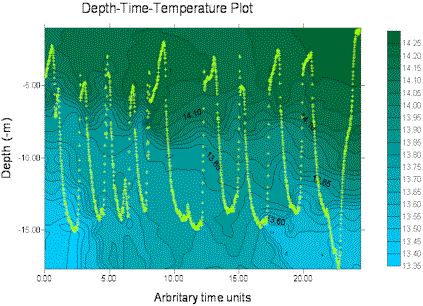
Figure 12. miniBAT profiles from offshore boat work
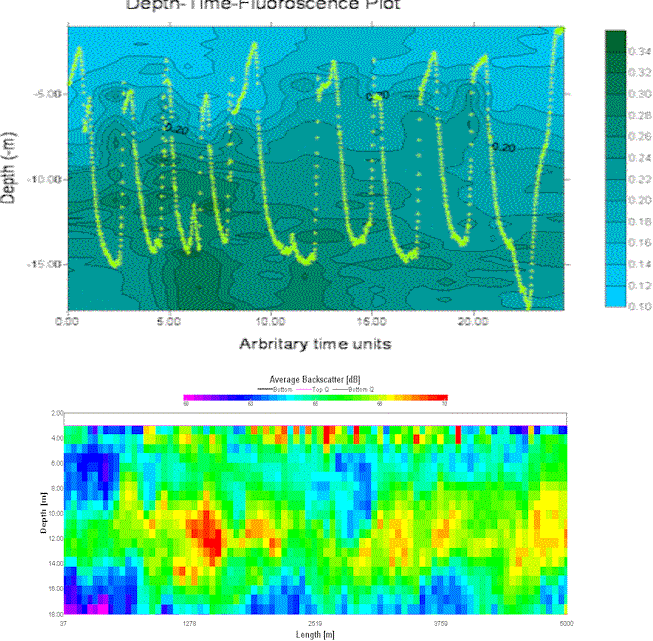
Figure 13. comparisons between miniBAT and ADCP data
Between site 3 (50°17.823’N, 004°23.950’W, 12:40 GMT) and site 4 (50º 20.778'N, 004º 21.186'W, 13:33 GMT) the CTD data shows the chlorophyll maximum being pushed upwards in the water column from a depth of 19m at site 3 to approximately 12m at site 4, despite the presence of an obvious thermocline to hold the phytoplankton in place at site 4 indicating that the phytoplankton are being pushed onshore. This could be occurring due to the south westerly winds in the area for at least a week prior to sampling, or due to the river plumes from the estuary encouraging onshore currents in Whitsand bay. The data displays how the physical mixing processes in the shallowing water push the phytoplankton and therefore the chlorophyll maximum closer to the surface.
Salinity decreases along the tow, moving inshore. There is little or no vertical stratification of salinity along the tow. Slightly lower salinity represents the present, but limited, influence of the river plume from the estuary mouth.
Temperature shows vertical stratification, and very slight horizontal stratification, maximum temperatures found at the surface and inshore. The thermocline is at approximately 5m depth at the beginning of the tow, and finishes at approximately 10m.
When comparing the miniBAT data and the corresponding part of the ADCP transect, done by scaling each data and displayed below, it was observed that the area of high fluorescence in the miniBAT data corresponds to an area of high intensity backscatter in the ADCP transect. The area of backscatter observed in the ADCP transect sits slightly higher in the water column than the area of fluorescence; it is probable that the backscatter is caused by zooplankton, as they sit higher in the water column than the phytoplankton seen in the miniBAT data and graze the phytoplankton downwards from above.All files relating to the minibat work, including grid files etc. can be found under group11\All work\28-06-04-off\Off_data\off_proc\minibat
Analysis of water samples - offshore work
A brief description for the procedure followed for measuring nitrate concentration is given in the directory:
group11\All work\28-06-04-off\Off data\off proc\chemanly
Profile shapes for chlorophyll concentration are approximately mirrored by those for silicon concentration (fig. 14). This indicates that the concentrations of chlorophyll and silicate could be linked by a biological process in the water column (primary production). As phytoplankton primary production takes place, the nutrients nitrate, phosphate (and silicate for diatoms) are absorbed. Hence, the water sample concentrations for these nutrients are seen to be low where chlorophyll concentration is high.
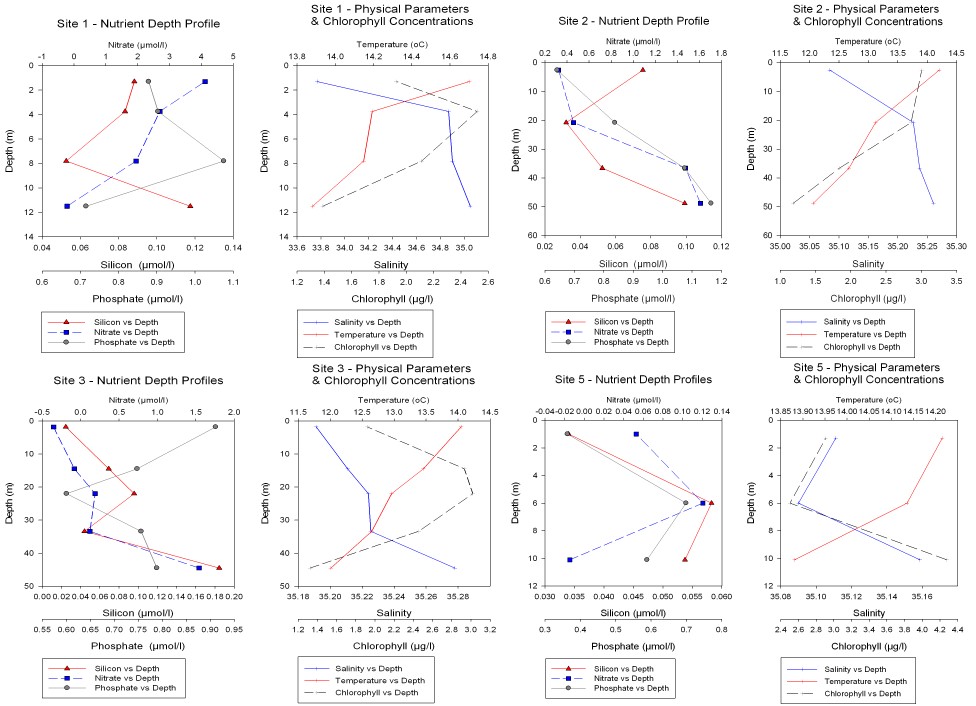
Figure 14. Nutrient concentration vs depth for stations 1,2,3 & 5
Chemical Analysis
Chlorophyll
Site 1 (behind breakwater: all lat. and long. positions of sites are given in table 2). There is a good correlation between depth of chlorophyll maximum and depth of nitrate and silicate minima at site 1 (behind the breakwater. Position of site in table 2).
Sites 2 and 3 (offshore) both show an opposing trend of high chlorophyll concentration at the same depth as low concentrations of silicate and nitrate (fig. 14). From data collected with the ADCP, site 2 (L4) appears to be located on a frontal system (fig. 11. Data file oadcp11003r.000 in the directory group11\all_work\28_06off\off_data\off_raw\adcp). The mixing conditions that produce frontal systems determine the availability of light and nutrients necessary for phytoplankton growth. (Pingree, 1975). This in turn has an effect on the depth of the chlorophyll maximum, and the concentration of chlorophyll present. At site 2 the chlorophyll maximum appears to be close to the surface. Surface concentration of chlorophyll at this site is high (3µg/l). The chlorophyll maximum has been pushed down from the surface waters to a depth of 20 m with concentrations of 3µg/l.
Site
5 (inshore). From the
CTD data the temperature and salinity profiles show that the water column is
well mixed and that the water depth is deeper than the thermocline depth. The
chlorophyll maximum has been pushed down to the bottom of the profile. The
absolute highest concentration of chlorophyll were recorded at the bottom of the
site 5 profile at 10m depth of 4.3µg/l. Site 5
was quite far inshore (table 2), and the presence of the chlorophyll maximum at depth
could represent phytoplankton being driven onshore from deeper offshore
thermoclines by currents and stormy weather.
Nitrate/nitrite
Brief analysis is given for sites where sufficient samples have been taken in order to provide a sensible interpretation
Site1 (behind breakwater). Nitrate and nitrite concentrations decrease with depth. However, concentrations of these nutrients do not increase as the Silicon levels do at 8m depth. This indicates that there is no bottom water nitrate and nitrite input. Since site 1 is landward of the breakwater, high nitrate and nitrite levels in surface waters may represent fresh water inputs, which are enriched by agricultural runoff. At site 1 we see the absolute highest concentrations of nitrate and nitrite from all the sites at 4µm/l at the surface.
Site
2 (offshore). Nitrate concentration (includes nitrite)
increases rapidly below 30m up to 1.7µmol/l. This indicates that there is little
or no mixing between the nutrient-rich bottom waters and the phytoplankton-rich
surface waters.
Site 5 (inshore). A good correlation of decreasing nutrient levels with increasing chlorophyll concentration exists, with the chlorophyll maximum and nutrient minimum at 6m depth.
Silicon
Site 1 (behind breakwater). The positioning of site 1 (see chart) behind the breakwater in the Sound has produced the expected results for chlorophyll with Silicon (fig. 14 - Station 1). These results are: a Silicon minimum corresponding to the depth of the chlorophyll maximum. Below the chlorophyll maximum concentration of Silicon increases.
Site 2 (offshore). Surface Silicon levels at site 2 are slightly higher than expected; this may be due to the end of the spring bloom of diatoms meaning that this nutrient is no longer scavenged in surface waters. Below the thermocline (see also CTD data in the directory: group11\all_work\28_06off\off_data\off_proc\ctdproc) the nutrient levels are much higher than the surface concentrations.
Site 3 (offshore). The absolute highest concentrations of Silicon were found in the deep (~45m) waters at this site, with values of 0.18µm/l.
Phosphate
Site 1 (behind breakwater). At this sampling station the phosphate concentrations do not follow the trend of silicon and nitrate. This anomaly may be due to the large fresh water input following a period of heavy rain, and also possibly the influence of the breakwater in close proximity.
Site 2 (offshore). At this site the highest (1.18µm/l) values of phosphate for all offshore sampling sites from the day were recorded, below the thermocline at around 50m depth.
Site
3 (offshore). the
phosphate levels are high at the surface and decrease with depth towards the
chlorophyll maximum (around 20m)
Dissolved
oxygen
Site
1 (behind
breakwater). The dissolved oxygen against
salinity plot shows that the water column is well mixed. Dissolved oxygen shows
a decreasing trend with increasing salinity up to 34.3 and then fluctuates with increasing salinity.
Site
2
(offshore).
Dissolved oxygen
decreases from 86% saturation to 73% saturation with increasing salinity. This is similar to site 3
(also offshore), at
which the saturation % decreases from 87% to 73%.
Biological Analysis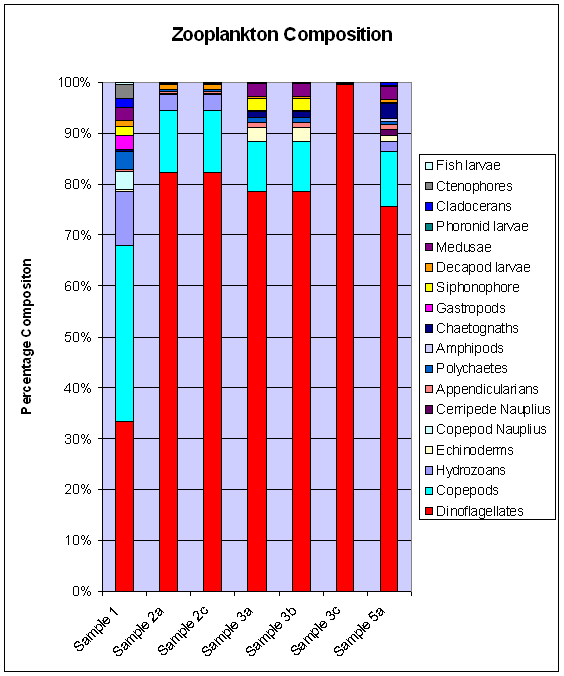
At all sites the dominant species are heterotrophic dinoflagellates, contributing to more than 70% of the population in all samples except for site 1 (behind the breakwater). At site 1, there are approximately equal numbers of copepods and heterotrophic dinoflagellates. This could be due to this site's position at the mouth of the estuary and hence higher phytoplankton abundance.
Sites 1 and 5 (both near shore sites)
contain the highest species diversity. This may be because of optimum physical
conditions and nutrient availability. From the data a dinoflagellate bloom was
observed at the inshore sites. These blooms are very important to the system due
to their contribution to annual primary production (Holligan et al. 1981). This
compliments the high zooplankton diversity seen at sites 1 and 5. Ctenophores
and fish larvae were only seen at site 1, again suggesting these sites' high productivity.
Site 3 shows a wide marine diversity, having large abundances of echinoderms, siphonophores and medusae (which the other sites were lacking or had much less of). Site 2, the L4 site, had comparatively low species diversity, having dinoflagellates as 80% of the community structure; a front was discovered in the ADCP data between sites 2 and 3. Fronts are significant in determining distributions in phytoplankton (Pingree et al. 1975).
Figure 15. Zooplankton composition from offshore samples
| Sample no. | Position | Location | Depth (m) |
| 1 | 50º 20.074'N 004º 08.135'W | site 1 | 14.2m to surface |
| 2a | 50º 15.163'N 004º 12.279'W | site 2 | 38m to surface |
| 2c | 50º 15.163'N 004º 12.279'W | site 2 | 50m to 38m |
| 3a | 50º 17.935'N 004º 22.618'W | site 3 | 23m to surface |
| 3b | 50º 17.935'N 004º 22.618'W | site 3 | 34m to 23m |
| 3c | 50º 17.935'N 004º 22.618'W | site 3 | approx 2m. Horizontal tow. 5 mins |
| 5a | 50º 17.214'N 004º 13.085'W | site 6 | 45m to surface |
Table 3. Positions and depths of zooplankton nets (200µm mesh). Offshore boat work.
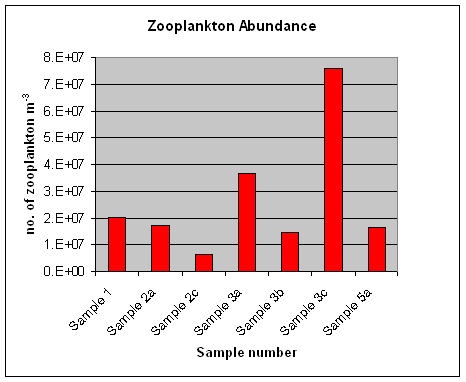
Figure 16. Abundance of zooplankton at the sites sampled during the offshore work. Note: sample 3c is of a horizontal tow at approx 2m depth.
This procedure aimed to develop an understanding of how the Tamar Estuary acts as a transition zone between the freshwater input and the coastal sea. Eleven stations were sampled on 02/07/04, shown in Figure 17, in conjunction with the ribs group (group 10) in order to produce a holistic view of the estuary. ADCP transects were used to distinguish velocity magnitude, shear direction and backscatter. CTD profiles were used to observe the features of the taken transect. Four Niskin bottles, attached to a CTD rosette, were used to gain water samples for nutrient, chlorophyll and dissolved oxygen analysis from varying depths. Plankton nets were towed at two sites, the most marine being site1 and the most freshwater, site 11, for direct comparison. A Secchi disc was used to determine the euphotic depth at most sites. Phytoplankton samples were taken from the surface and preserved in Lugols iodine solution for laboratory analysis.
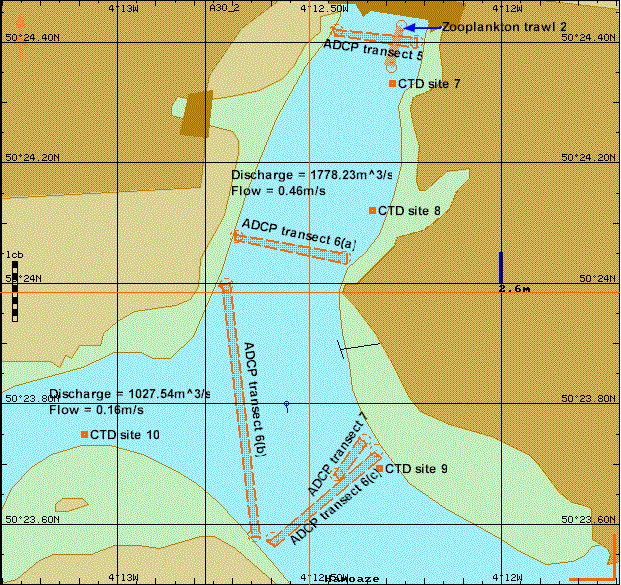
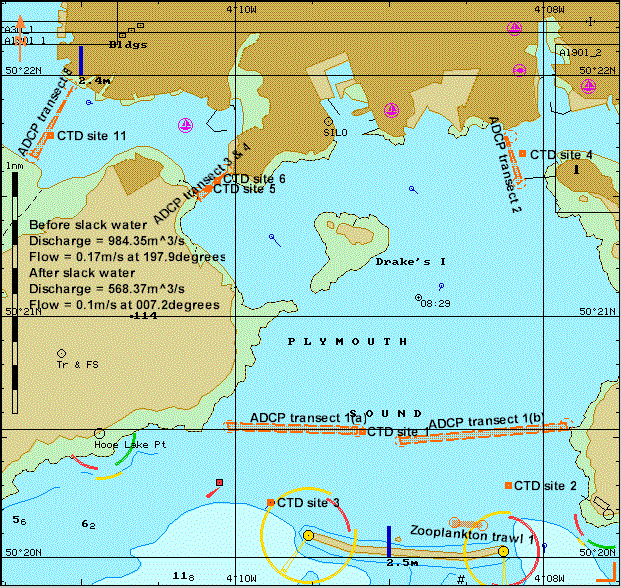
Figure 17. Sites sampled during estuarine boat work on Bill Conway
Temperature and salinity depth profiles
Site 1 – In the middle of the Sound, the water column is fairly well mixed, with very little difference between the surface and bottom water temperature and salinity.
Site 2 – On the East of the sound, there is stratification in the surface waters, with a thermocline at 6m depth.
Site 3 – West of Sound. Still fairly well mixed with a thermocline at 6m.
Site 4 - Cattewater – Gradual decrease in temperature from the surface (15.3 °C) to 14. 8 °C at 6m. Surface salinity is lower at 32.6 compared to previous sites, and increases with depth i.e. there is a small salt wedge.
Site 5 & 6 –
Site 7 –
Site 8 – The observed front by the Tamar Bridge. It is more saline at the surface and increases with depth to 33.5. There is a relatively large change in temperature between warm surface waters and colder bottom waters.
Site 9 – The observed front by the Lynher – This profile shows a split thermocline with well mixed bottom waters.
Site 10 – Mouth of Lynher – More saline and colder at the bottom. Temperature at the surface is 16.35 °C
Site 11 –
ADCP ANALYSIS-
Bill Conway,
|
ADCP transect- Time (GMT) |
GPS position |
Location |
Tidal state |
Description/Analysis, from Magnitude velocity and Average backscatter
profiles |
|
Transect
1 START;08.42 END;09.11 |
Start;
50˚20.531N
004˚10.030W Finish;
50˚
20.546N
004˚ 07.826W |
Transect from East to
West across the width of |
Ebbing |
Velocity magnitude is homogenous throughout the transect. Max velocity reaching 0.4 m/s. Relatively clear water column represented by little backscatter. Intermittent area of high backscatter at surface down to around 4m. Turbulence caused as sea bed shallows, around 6.9m. |
|
Transect
2 START;10.20 END;10.23 |
Start; 50°
21.559N 004°08.159W Finish; 50°
21.741N 004°
08.216W |
Across width of
Cattewater. Top of |
Ebbing |
Again velocity is relatively homogenous with peak velocity at 0.4 m/s. Slower velocities dominate away from the start of the transect (further south). Sporadic backscatter at 74 db caused by extensive mooring facilities. Water column clear from 1.5m -6m. |
|
Transect
3 START;10.52 END;10.56 |
Start;
50°
21.598N 004°
10.074W Finish;
50°
21.512N 004°
10.237W |
Width of the narrows |
Ebbing- just before slack |
Average
velocity magnitude maintained around 0.4 m/s, although velocity increases
through the |
|
Transect
4 START;11.46 END;11.49 |
Start; 50°
21.615N 004°
10.066W Finish; 50°
21.527N 004°
10.234W |
Width of the narrows |
Flooding |
Similar transect to above. Higher velocities found at deeper waters in middle of the channel. Due to the tidal state the higher velocities may be caused by the incoming water. Consistent layer of backscatter from surface to around 5m depth, turbulence and air bubbles present from tug and passing powerboat |
|
Transect
5 START;12.32 END;12.34 |
Start;
50°
24.427N 004°
12.321W Finish;
50°
24.425N 004°
12.111W |
Approx. parallel to Tamar and Royal Albert bridge |
Flooding |
This transect was furthest upstream out of all the transects carried out. Again maximum velocity was found at the deepest region of the transect. The majority of the transect demonstrates a relatively turbid water column, with backscatter at 72db. An area of strong backscatter was found in the same region as greatest velocity magnitude, suggesting sediment disruption on the river bed. The water is approximately 3m deep at the outside of the channel; backscatter here suggests small communities of plankton. |
|
Transect
6a START;13.17 END;13.21 |
Start; 50°
24.004N 004°
12.314W Finish; 50°
23.987N 004° 12.649W |
Width of Tamar just before confluence with the Lynher |
Flooding |
Velocities similar to site 5, homogenous throughout the transect. Reaching 0.4 -0.5 m/s. Strong backscatter suggests disrupted sediment caused purely by the velocity as well as increased turbidity from upstream. |
|
Transect
6b START;13.22 END;13.27 |
Start; 50°
23.977N 004°
12.657W Finish;
50°
23.587N 004°
12.611W |
Across Lynher just before confluence with the Tamar |
Flooding |
The majority of the channel has a velocity magnitude of around 0.3 m/s, until the Tamar joins with the Lynher, flow velocity increases at the Eastern end of the transect. Strong backscatter represents turbid waters and presence of plankton from the Tamar. |
|
Transect
6c START;13.28 END;13.31 |
Start;
50°
23.582N 004°
12.605W Finish; 50
23.744N 004
12.247W |
Width of Hamoaze after confluence of Lynher and Tamar |
Flooding |
Rapid change in velocity magnitude from 0.3m/s to around 0.75m/s. A clear front demonstrating this is seen, faster velocities undercut the slower rates, suggesting a denser water mass. Where the front can be seen there is a layer of strong backscatter reaching a depth of 5.5m, representing either plankton communities gathered here and scum build up where the two masses meet. Strong backscatter from the river bed may be present due to potential emissions from two local inflow pipes from Kinterbury point. |
|
Transect
7 START;13.40 END13.42 |
Start; 50°
23.724N 004°
12.332W Finish; 50°
23.689N 004°
12.412W |
Short transect just above transect 6c. Eastern side of river channel |
Flooding |
Again rapid change in velocity magnitude demonstrates the front that was physically seen. Undercutting by faster velocities is evident. Strong backscatter is evident along the western end of the transect with an underlying layer of similar strength (75db) throughout the transect. The mass of water with a slower velocity seems to be harboring more turbid water, and vertical mixing and the corresponding turbulence will also cause the strong backscatter. |
|
Transect
8 START;14.18 FINISH;14.21 |
Start;
50°
21.953N 004°
11.055W Finish; 50°
21.657N 004°
11.330W |
Transect parallel to |
Flooding |
Again velocity variations correlate with the trends identified at transects 6 and 7. Velocity magnitude reaches 0.9 m/s around the southern point of the transect, shallow waters cause the velocity to decrease. A stretch of backscatter of 75db runs along two thirds of the transect reaching a depth of around 6m. Suggesting the presence of plankton throughout the photic layer. |
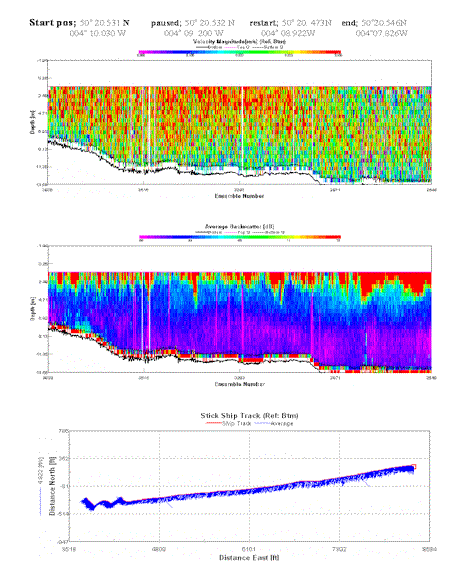
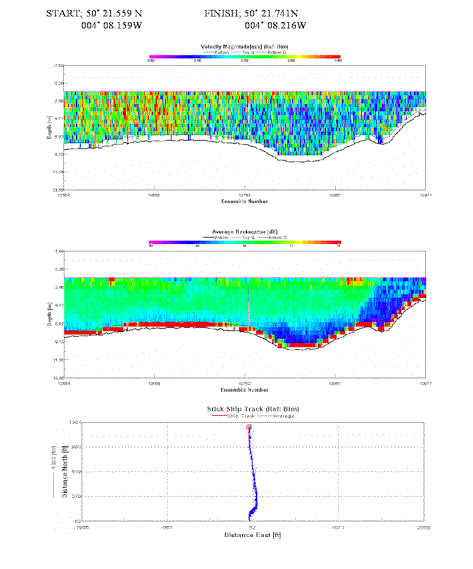
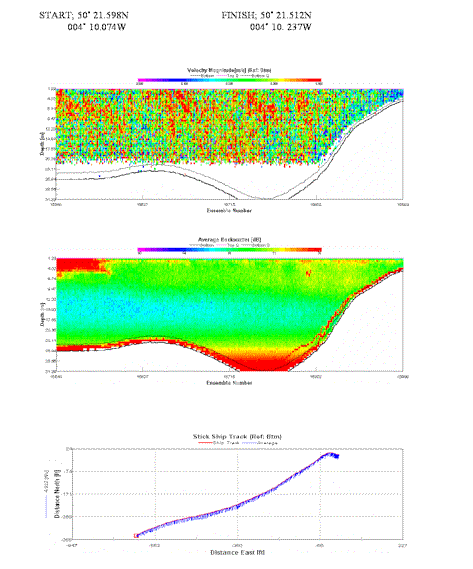
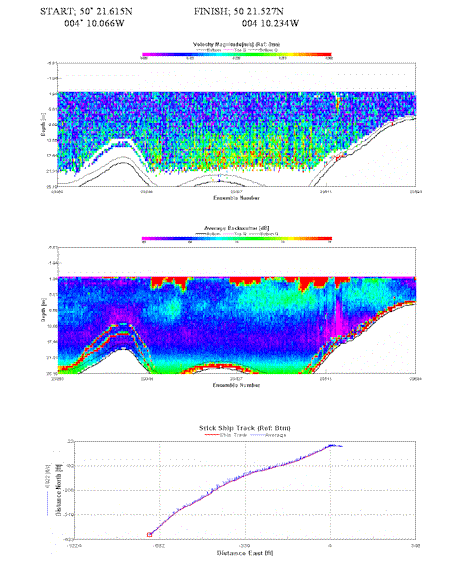
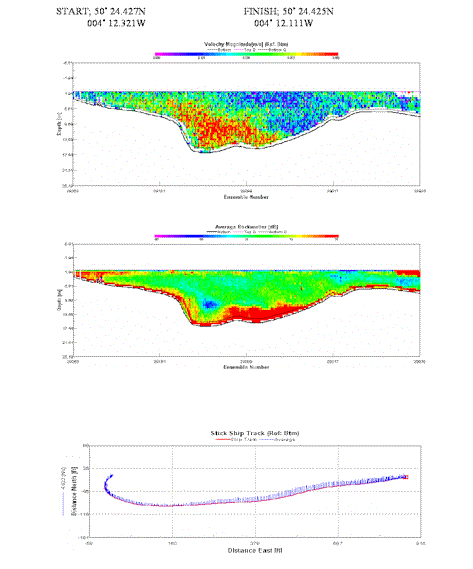
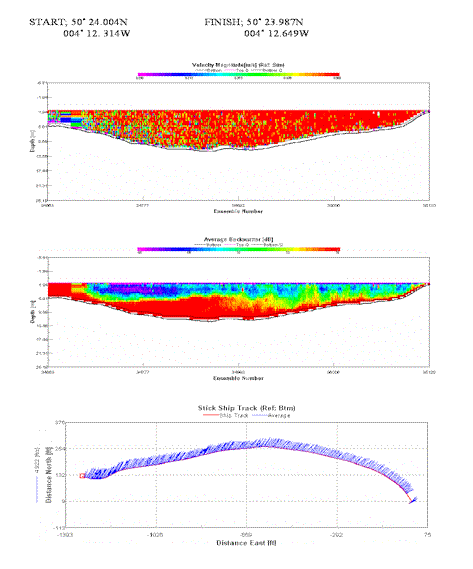
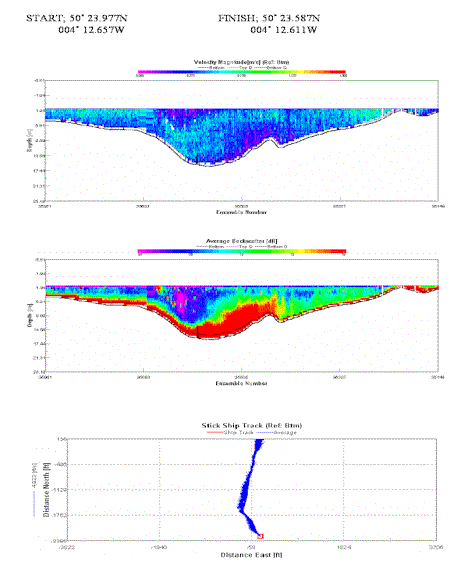
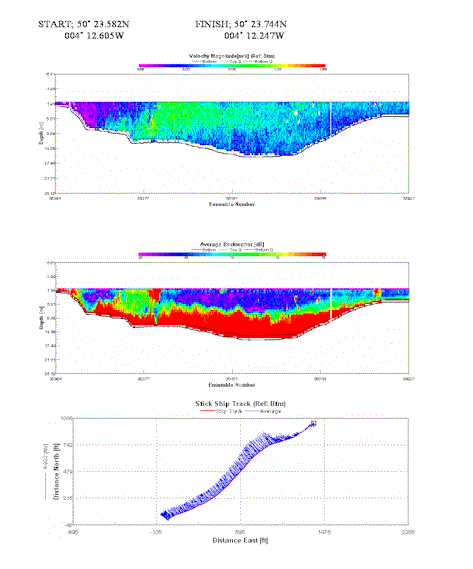
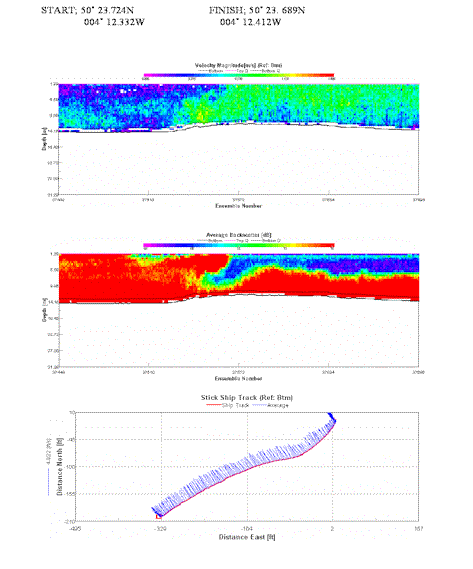
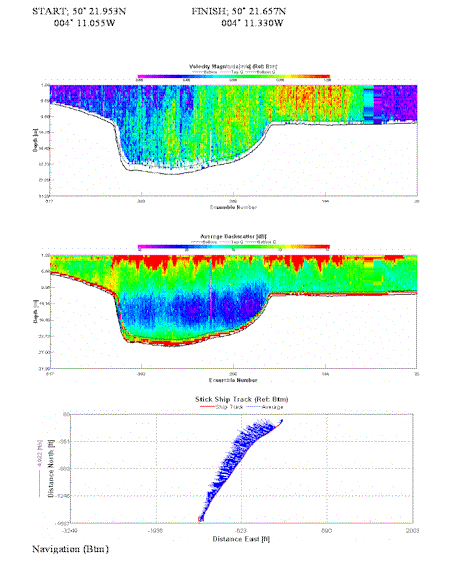
Table 4. Analysis of estuarine ADCP data
Secchi disc analysis
Site 1 has the greatest euphotic zone at 12.18m because it is the most marine environment and has the least suspended sediment, due to an increased amount of Na+ ions. These tend to cause sediment particles in the water column to coagulate; falling to the bottom.
At site 2 colour filters were used, to show which wavelengths of light were attenuated first. The first wavelength to be completely absorbed at 1.21m was red, followed by green at 1.4m then blue at 1.88m. This indicates a high phytoplankton abundance at this site, as a large amount of green light would be absorbed due to the high chlorophyll concentration.
After site 2 the euphotic zone decreases in depth with increasing salinity. At site 7 by the Tamar bridge, the euphotic depth is 10m higher in the water column than at marine site 1. This is because of increased suspended particulate matter (SPM) from the riverine input, which increases turbidity. Filters used at this site show that there is not as much phytoplankton, since the green wavelengths penetrated deeper into water column.Nutrient concentrations with depth profile analysis.
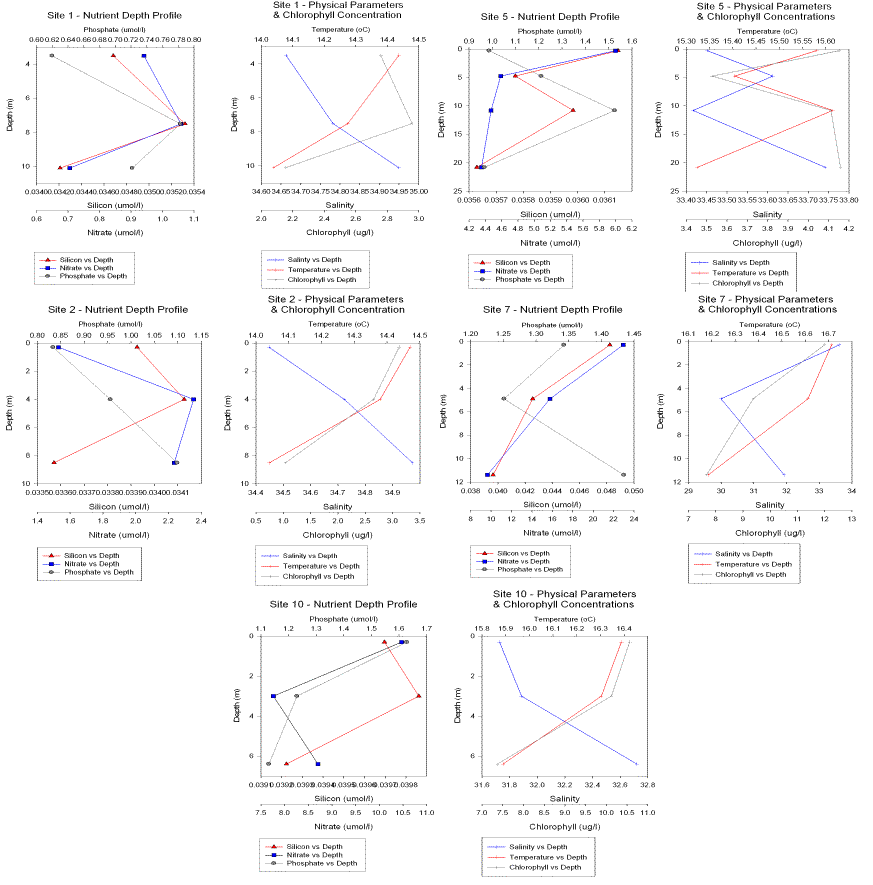
Figure 18. Nutrient and chlorophyll profiles from 02/07/04.
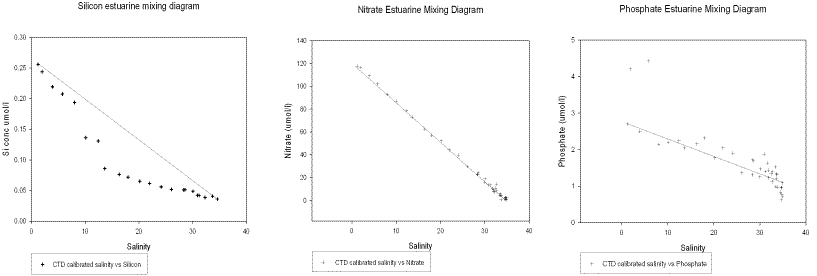
Figure 19. Estuarine mixing diagrams from boat work carried out on 02/07/04
Chlorophyll
Site
1. By the
breakwater. The chlorophyll concentrations in this profile are the lowest of
this set of profiles from
Site
2. Towards the east
of the Sound. The maximum chlorophyll concentration occurs at the top of this
profile in the surface waters of 13.2μg/l.
Site
4. This profile
shows opposing chlorophyll and nutrient concentrations, as phytoplankton
abundance increases with depth
Site
5.
The Narrows.
This profile was taken at slack water in a region where there is high velocity
flow over much of the tidal cycle. The
nutrient and chlorophyll depth profiles show extreme mixing, with fluctuating
peaks of high and low concentrations.
Site
7.
By the Tamar
bridge. The surface concentrations of chlorophyll, nitrate and silica are all
very high (12μg/l, 22μmol/l and 0.048μmol/l respectively) and
decrease with depth.
Site
10. Mouth of the
river Lynher. At this site fresh
water anomalies were observed from the boat and the definition between the fresh
input and the rest of the water mass could clearly be seen as a foamy, bubbly
film which was caught in the convergence between water masses.( See
adjacent photo.)
See
adjacent photo.)
Elevated
silicate concentrations may represent the presence of a diatom population around
this front or simply high surface nutrient concentration. The chlorophyll
concentrations at this and other upstream sites may be elevated due to there
being less fresh water zooplankton than saltwater zooplankton at the sites
towards the seaward end of the estuary. The maximum chlorophyll abundances will
appear closer to the surface the further up the estuary as high suspended
sediment load in the water column reduces the euphotic depth.
Nitrate/nitrite
Site
1- In the middle
of the Sound we see the lowest nitrite and nitrate concentrations of only 0.93
μmol/l in the surface waters, peaking in this depth profile at around 8m
with 1.08μmol/l and almost depleted in the bottom waters. This indicates
that the supply of nitrate and nitrite to Plymouth sound is not from the saline
bottom waters but from the fresher riverine waters.
Site
2 The nutrient
concentrations are still relatively low and deplete towards the bottom of the
profile with a peak at 4m depth.
Site
4 the nitrate
concentrations decrease with depth. The
surface nitrite and nitrate concentration at this site is elevated relative to
other sites with a maximum of 14.5μmol/l. This elevation suggests a
localised input of nitrogen to the estuary. Looking at the charts this location,
at the mouth of the dredged Cattewater river, is a heavily polluted input into
the sound with silos and other sewage pipes discharging organic and inorganic
nitrogen forms. There is an Esso wharf also located by the site.
Site 5
- Nitrate concentration generally decreases with depth.
Site
7 By the Tamar
bridge. The surface concentrations of chlorophyll, nitrate and silica are all
very high (12μg/l, 22μmol/l and 0.048μmol/l respectively) and
decrease with depth. At this site inputs upstream may be affecting the
concentrations measured. On the charts, sluices and silos as well as other pipes
all discharge into the river. There is also agricultural land further upstream,
and runoff from these lands (pesticides. herbicides and excretory products) may
also be contributing.
Site
10 The water
profiles show high surface nitrate concentration (10μmol/l) which decreases
with depth (however are still relatively high at the bottom of the profile at 8
μmol/l)
Silicon
Site
1. The distribution
of Silicon and chlorophyll follows the same pattern as nitrate, with peaks of
0.0352μmol/l and 2.85μg/l respectively at 8m depth.
Site
2. The nutrient
concentrations are still relatively low and deplete towards the bottom of the
profile, with a peak at 4m depth.
Site
4.
The silicon
concentrations decrease with depth.
Site
5.
Chlorophyll and
silica concentrations peak at 10 m depth.
Site
7. By the Tamar
bridge. The surface concentrations of chlorophyll, nitrate and silica are all
very high (12μg/l, 22μmol/l and 0.048μmol/l respectively) and
decrease with depth.
Site
10.
Surface
chlorophyll and silica concentrations are also high (10.5μg/l and 0.0397μmol/l)
and follow the same trend as nitrate of decreasing concentration with depth.
Phosphate
Site 1. The lowest estuarine phosphate levels were observed here, being 0.62μmol/l at the surface and increasing in concentration with depth.
Site 2. Very low phosphate concentration at the surface, increasing with depth.
Site 5. At the narrows the water column is well mixed, with high concentrations of around 1.5μmol/l.
Site 7. High surface concentrations (1.35μmol/l) increasing with depth to 1.43μmol/l.
Site 10 (Tamar Bridge). The highest estuarine concentrations were measured here, being 1.6μmol/l in the surface waters. The elevated phosphate levels at these upstream sites are possibly due to agricultural runoff (organo-phosphates from fertilisers). The general trend of estuarine phosphate was found to be that surface concentrations increase the further upstream.
Dissolved
oxygen
Geophysics boat
practical
The aim of the
geophysical practical was to conduct a detailed seabed survey of the region to
the south of
From analysing
the data it is possible to calculate, using Pythagoras theorem, the distance
away from the centre of the trap objects are and also the height of objects
protruding from the seabed.
Multiple
sites were surveyed to try and find some interesting data, but due to the divers
over the James Egan Layne it was not possible to get as much information as
desired. The analysis below refers to site 3 of the sidescan sonar sites.
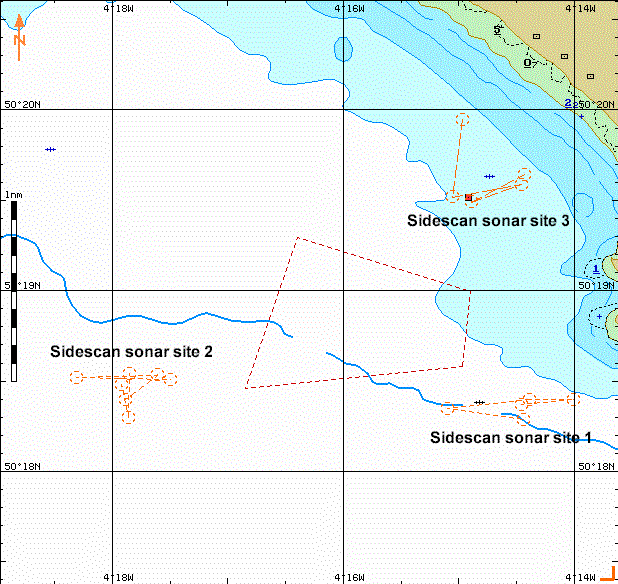


Fig. 20 All sidescan survey sites Fig. 21 sidescan survey site 3 Fig. 22 sidescan sonar plot Fig. 23 vertical rock strata
(group11\all_work\05-07gp\pics\geopic)
Analysis
Summary
The data collected from
From the samples of phytoplankton and zooplankton collected from various parts of the estuary and offshore, we concluded that the tail-end of a dinoflagellate bloom had been sampled offshore. There is a higher species diversity of zooplankton in the marine environment than in the estuary. High quantities of freshwater phytoplankton were collected. Chlorophyll data and plots complement these findings.
CTD profiles show the development of a seasonal thermocline offshore. The riverine influence in terms of freshwater is not easily seen beyond the breakwater at the southern end of Plymouth Sound. In the estuary, fresh water flows seawards overlying the more saline. Since the estuary is partially mixed, the tidal influence can be seen up the estuary to Calstock, where 0 salinity fresh water overlies water of salinity ~7.5.
An appreciation
of the use of sidescan sonar in geophysical seabed surveying, was also achieved.
Grabemann, I., Uncles, R. J., Krause, G. and Stephens, J. A. (1997) Behaviour of Turbidity Maxima in the Tamar (U.K.) and Weser (F.R.G.) Estuaries. Estuarine, Coastal and Shelf Science 45:235-246.
Holligan, P. M., Viollier, M., Dupouy, C. and Aiken, J. (1983) Satellite studies on the distributions of chlorophyll and dinoflagellate blooms in the western English Channel. Continental Shelf Research 2(Nos 2/3):81-96.
Morris, A. W., Bale, A. J. and Howland, R. J. (1981) Nutrient Distributions in an Estuary: Evidence of Chemical Precipitation of dissolved Silicate and Phosphate. Estuarine, Coastal and Shelf Science 12:205-216.
Pingree, R. D. (1975) The advance and retreat of the thermocline on the continental shelf. J. Mar. Biol. Ass. UK 55: 965-974.
Pingree, R. D., Pugh, P. R., Holligan, P. M. and Forster, G. R. (1975) Summer phytoplankton blooms and red tides along tidal fronts in the approaches to the English Channel. Nature 258:672-677.