Life-Cycle of Emiliania huxleyi
John Green
Plymouth Marine Laboratory
Citadel Hill
Plymouth PL1 2PB, UK.
Email: JGR@wpo.nerc.ac.uk
Sexual reproduction in all species of plants and animals is characterised by
the fusion of nuclei from specialised cells (gametes - e.g. sperm and ova in
mammals), derived from the parental organisms. The cell that results from
the fusion of two gametes develops subsequently to form a new individual.
The parental cells are diploid, having twice as much DNA as the gametes,
which are haploid. In many organisms, the haploid and diploid stages of
the life-cycle are different in form. Thus, in flowering plants, the male
haploid stage consists of the pollen grain, and the female haploid stage
consists of the embryo sac. All other parts of the familiar flower-bearing
plant are diploid, and this diploid stage is the only stage that can lead an
independent existence. However, within the algae, haploid and diploid
generations may be equally well-developed and able to exist independently,
each reproducing vegetatively (i.e. the clonal offspring having identical
DNA, and the same amount of DNA as the parent). In such cases, the haploid
and diploid stages may be similar or different in form.
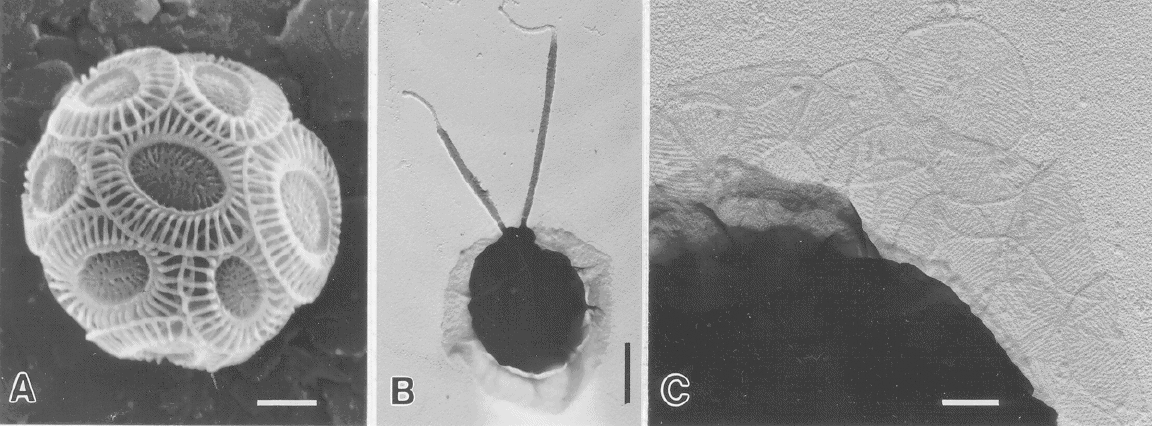
It has been known for some time that the the life-cycle of Emiliania
huxleyi involves several cell types (Klaveness, 1972) that can maintain
themselves by vegetative reproduction. They include the non-motile
coccolith-bearing cell (the C-cell; Fig. A), naked non-motile cells
(N-cells), and motile scale-bearing swarmers (S-cells; Figs B, C). How they
are related genetically is not certain, though early evidence suggested
that the S-cells may be haploid relative to the C-cell (Klaveness,
1972).
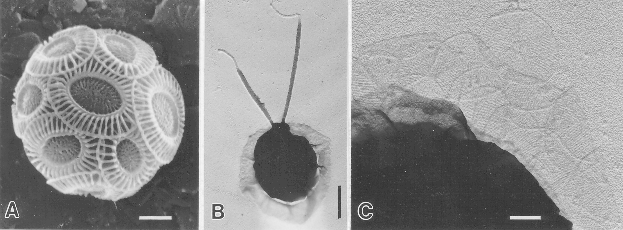
(A): non-motile C-cell of Ehuxleyi showing the investment of
overlapping coccoliths (scale bar = 1.0 micron); (B):
scale-bearing cell (S-cell) of Ehuxleyi (scale bar = 1.0 micron);
(C): body-scales from an S-cell of Ehuxleyi (scale bar = 0.2
micron). (SEMs from John Green)
Recent work has concentrated on elucidating this problem by measuring
relative levels of DNA using flow cytometric analysis of
fluorochrome-stained DNA in the different cell types (Green et al., 1996).
In this technique, the flow cytometer measures the fluorescence produced by
the stained DNA in individual cells when illuminated using light at
particular wave-lengths. The fluorescence produced is proportional to the
DNA-content of the cells. It was successfully demonstrated that the motile
S-cells have a DNA level half that of the coccolith-bearing cells and are,
therefore, haploid relative to the C-cells (Fig. D). This strongly suggests
that Ehuxleyi does indeed have a life-cycle involving a sexual stage with
the S-cells acting as gametes, though they themselves can reproduce
vegetatively. However, fusion of gametes as part of sexual reproduction has
not yet been observed. It is also important to determine what initiates the
changes between the various cell forms; for example, the N-cells and S-cells
may arise spontaneously from C-cells in culture, but the exact triggering
conditions are not yet known. Such information is urgently needed in order
to understand the way that the Ehuxleyi life-cycle operates in the
oceans, and in understanding how and why blooms of this widespread and
cosmopolitan organism develop.
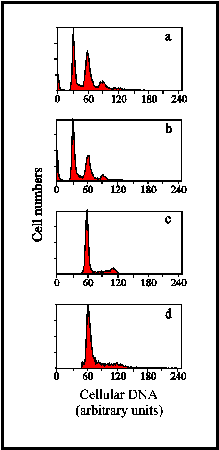
(D): Relative DNA content of Ehuxleyi cells. (a):
peaks from a culture containing C-, N- and S-cells. The principal peaks are
at 30 and 60 units of DNA, with smaller peaks at 90 and 120.
(b): in a culture containing principally S-cells, the main
peaks are again at 30 and 60 units of DNA. (c): in a culture
of C-cells only, the principal peaks are at 60 and 120 units.
(d): this culture, used as a reference clone, also contained
only C-cells with the main peak at 60 units and a small peak at 120 units.
It is presumed that 60 units represents a diploid complement of DNA for one
cell; 30 units indicates a gamete and 120 units comes from a cell just
before binary fission, or just before it releases gametes. In addition, 60
units could also represent a gamete about to divide to form 2 gametes.
(Diagram by Glen Tarran)
For further details of the work above, see Green et al. (1996) and for
preliminary alternative explanations of the DNA complements in different
Ehuxleyi cell types, see Green et al. (1996) and Medlin et al. (1996).
For a general discussion on life-cycles within the algal division
Haptophyta, see Billard (1994).
References
Ehux
home page