"Geological Context" of Emiliania huxleyi
Patrizia Ziveri
Geomarine Center
Vrije Universiteit Amsterdam
de Boelelaan, 1085
1081HV Amsterdam
The Netherlands
Email: zivp@geo.vu.nl
Emiliania huxleyi was first identified by Lohmann (1902) using a
light microscope. The details and beauty of its structure were not visible
however until the development of electron microscopes. The structure was
then described from some of the earliest electron microscope observations
of plankton - Braarud et al (1952), Deflandre & Fert (1954), Black & Barnes
(1961). The phylogenetic origins of Ehux are well established from
the fossil record (Perch-Nielsen, 1985; Romein, 1979; Gallagher, 1989;
Young et al., 1992). The genus is descended from other coccolithophore
types, some of which are still present in today's ocean. It is believed to
have evolved from Toweius through Reticulofenestra and
Gephyrocapsa (Young, 1989; Young et al., 1992).
Ehux is only seen in relatively young sediments, and must have
evolved fairly recently. The first appearance datum (FAD) of Ehux in
the sediment is used as a biostratigraphic marker. Depending on the
different proxies used and the location, this entry has been dated at
~270,000 years ago using correlation with planktic foraminiferal delta-O18
records (Gartner and Emiliani, 1976), at ~268,000 years ago late in oxygen
isotopic stage 8 (Thierstein et al., 1977) and at ~285,000 years ago (Ahagon
et al., 1993). Using correlations based on magnetostratigraphy, Weaver
(1993) dated it at ~242-268,000 years ago, while Hills and Thierstein (1989)
estimated ~230,000 years ago.
In the living coccolithophorid flora, Ehux has the widest
distribution and largely dominates the living assemblages world-wide with
exception of a few oceanographic regimes. It also forms gigantic blooms
readily visualised by satellite imagery. This blooming life-style started
between ~80,000 and 50,000 years ago, marking the beginning of the so called
Ehux Acme zone in the fossil record. This Acme zone starts later as
one progresses towards the poles. The earliest onset time has been reported
from the Tropics (McIntyre 1967, Thierstein et al., 1977, Jordan et al.,
1996). In the Antarctic Ehux is absent and in the Arctic the acme
zone is terminated at the start of the Holocene. In shallow marginal seas
and inland seas, and in upwelling areas the Ehux Acme Zone began
later than in other parts of the ocean or is absent. In these areas
Gephyrocapsa spp., mainly G. oceanica, are generally more
dominant (Okada and Honjo, 1975; Ziveri and Thunell, 1995). Ehux may
have taken over an ecological niche formerly dominated by Gephyrocapsa
spp.

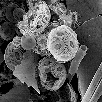
Two SEM images showing Ehux coccoliths and whole coccospheres
(some degraded) caught in a sediment trap (58 29'N, 20 29'W, 1044m depth) in
the northeast Atlantic. The sediment trap interrupted the transit of these
particles from the surface water photic zone (where the cells grew) to the
sea floor (where they would have been incorporated into marine rocks, if
they escaped dissolution). (Photos from Patrizia Ziveri and Saskia Kars).
[click on the small pictures to view them at full size]
Coccolithophores are a major contributor to the carbonate in deep sea
sediments. In the Northeast Atlantic during the last glacial-interglacial
cycle, coccoliths comprised 70-80% of the total carbonate during
interglacial times, and a variable but lower percentage during glacial times
(van Kreveld et al., 1996; Lotoskaya and Ziveri, 1995; Lotoskaya et al., in
prep.). Discrepancies exist between the estimation of export production and
the estimation of sedimentary preservation of calcium carbonate from
coccoliths of Ehux. It is still unknown how much of the calcite
produced during bloom events is stored in the sediment record, and how much
is dissolved back into the water. The geological branch of the
Global Emiliania Modelling (GEM) initiative
is working in great detail on this problem. Using sediment trap techniques
(capable of intercepting particle fluxes continuosly, in time fractionated
sequences and over a long period of time) we are able to quantify the
carbonate coccolithophore export production, and then compare it with the
rate of accumulation of coccolithophore assemblages in surface and in deeper
sediments. It has been shown that Ehux largely dominate the total
coccolith flux in the northeast Atlantic (Samtleben and Bickert, 1990;
Knappertsbusch and Brummer, 1995; Ziveri et al., 1996; Ziveri and Broerse,
1996), in the Northeast Pacific (Ziveri et al., 1995) and in the eastern
Mediterranean (Ziveri et al., 1995). Unfortunately, only very few
coccolithophore flux data and coccolith accumulation rates are available.
References
Ehux
home page

