Global Distribution of Coccolithophore Blooms
Christopher W. Brown
NOAA/NESDIS/ORA E/RA3
Washington, DC 20233
Email: chrisb@orbit.nesdis.noaa.gov
Blooms of the coccolithophore Emiliania huxleyi regionally act as
an important source of dimethyl sulfide (DMS) and calcium carbonate, and alter
the optical properties of the surface mixed layer (Balch et al., 1991;
Holligan and Balch, 1991). These blooms, often covering vast areas, can be
identified in visible satellite imagery because of the large amount of light
backscattered from the water column. Their presence gives the ocean a milky
white or turquoise appearance. The ability to detect E. huxleyi
blooms in satellite imagery, in addition to furnishing biogeographical knowledge of
the species at time and space scales unattainable with shipboard sampling,
provides a method to assess their biogeochemical importance on basin to
global scales.
Global composites of Coastal Zone Color Scanner (CZCS) imagery (Feldman
et al., 1989) were used to map the distribution pattern of E. huxleyi
blooms
and to estimate the magnitude and periodicity of their CaCO3 and DMS
production in the world's oceans (Brown and Yoder, 1994). Pixels of 5-day
composite imagery from the entire CZCS mission (November 1978 to June 1986)
were classified into either bloom or non-bloom classes based on their mean
normalized water-leaving radiances using a supervised, multispectral scheme.
This empirically-based classification technique is common in terrestrial
remote sensing, but has only recently been applied by oceanographers. A
classification algorithm was developed which compared the spectral signature
of known E. huxleyi blooms (e.g. (Holligan et al., 1983) to spectral
signatures of non-bloom conditions. Spectral signatures of E. huxleyi
blooms, "clear" blue water, sediment-laden water, "whiting" (suspended lime
muds), and atmospheric haze were extracted from CZCS imagery. Decision
boundary values for each of five spectral feature characters were assigned
that would allow the blooms to be spectrally distinguished from the other
conditions. An independent data set was also used to establish that the
algorithm was effective in distinguishing coccolithophore blooms from the
other water conditions, with the exception of whitings, at the spatial
resolution of the global imagery. The classified images generated from the
scheme were then combined into monthly, annual and mission climatologies of
bloom and non-bloom locations.
Spectral signatures similar to that of E. huxleyi blooms were
found to be most extensive at subpolar latitudes, particularly in surface
waters of the North Atlantic, the North Pacific and the Argentine shelf and
slope. (Plate 1). Classified blooms covered an average of 1.4 x 106 km2
annually, with the subpolar latitudes accounting for 71% of this area. The
classified blooms at these higher latitudes were inferred to represent the
presence of E. huxleyi blooms because the classification scheme
proved efficient in these regions and their locations are supported by
previous biogeographic investigations. Numerous classified blooms, often
quite extensive, were also detected at low latitude marginal seas, though
the conditions responsible for this signal are equivocal. Seasonally, the
classified blooms in subpolar oceanic regions achieved their greatest
spatial extent in summer to early autumn, while those in lower latitudes
peaked in mid winter to early spring.
Two important caveats of this approach should be noted. First, the
results displayed in Plate 1 reflect the distribution pattern of
coccolithophore blooms occurring in the surface layer and are biased toward
the declining stage (stationary phase) of the bloom. Detection of blooms is
sensitive to light backscattered from approximately one attenuation depth
and is primarily a function of detached coccolith concentrations. Blooms
composed primary of cells or occurring at depths deeper than that sensed by
the CZCS would be missed. Second, the distribution pattern of blooms and
their spatial extent are biased by both image coverage and regional
atmospheric conditions.
The amount of calcite-carbon and DMS-sulfur produced by the classified
E. huxleyi blooms was estimated using the mean annual areal extent of
the
blooms and representative values of mixed layer depth, average cell
concentrations found in blooms, DMSP concentration per cell, and mass of
calcite per coccolith. The blooms detected at subpolar latitudes (40ˇ -
60ˇ) are estimated to produce an average of 0.4 - 1.3 x 106 metric ton
CaCO3-C and 10,000 ton DMS-S annually. These standing stock estimates
suggest that satellite-detected blooms play only a minor role in the annual
production calcite and DMS on a global scale.
Although no satellite ocean color sensor has operated since the demise
of the CZCS in June 1986, future missions, such as the Sea-viewing
Wide-Field-of-View Sensor (SeaWiFS), will allow E. huxleyi blooms to
be monitored once again. These dedicated ocean color missions, in conjunction
with techniques to estimate coccolith and cell concentrations in
coccolithophore blooms from satellite imagery (Ackleson et al., 1994), will
improve our ability to assess the impact of coccolithophore blooms on the
carbon and sulfur cycles in the future.
References
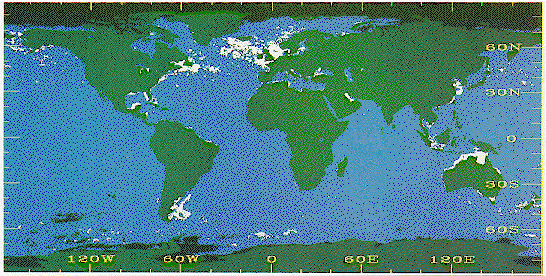
Plate 1. Mission climatology of classified coccolithophore blooms in the
world's oceans. The maximum spatial extent of blooms detected during
this period are displayed. Coccolithophore bloom class = white, non-
bloom class = blue, land = green, black = lack of data. (From Brown and
Yoder, 1994)
[This page is reproduced with permission from an article in Oceanography
8(2):59-60]
SeaWiFS images are now also being analysed to determine the global
distribution of coccolithophore blooms. Click
here
to see.
Ehux
home page